- Estructura de las paredes del laberinto membranoso.
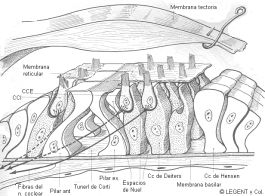
- Histología del órgano de Corti.
- Distribución de la inervación de las células ciliadas.
 HISTOLOGÍA DEL ÓRGANO DE CORTI.
HISTOLOGÍA DEL ÓRGANO DE CORTI. - Células de sostén.
Están fijas sobre la membrana basilar, dan soporte a las cc sensoriales y son de tres tipos: las que forman los pilares, las de sostén de las CCE y las de sostén de las CCI.
Tienen varias funciones:
- Dan soporte mecánico y metabólico a las cc sensoriales.
- Separan los compartimientos que contienen endolinfa de aquellos con perilinfa.
- Algunas participan en la formación y mantenimiento de ambos fluidos.
En su estructura presentan especializaciones de membrana, uniones intercelulares, tales como las uniones de hendidura o tipo "gap" (gap junctions), desmosomas (desmosomes), uniones adherentes (adherens junctions) y uniones estrechas (tight junctions).
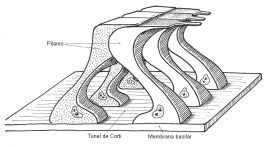 Están situadas en la parte media del órgano de Corti y son de forma piramidal y son un tanto particulares o propias de esta zona. Los dos pilares, externo e interno, formados por estas cc, delimitan junto con la membrana basilar el túnel de Corti. Están situadas en la parte media del órgano de Corti y son de forma piramidal. Están dispuestas en dos filas, externa e interna, formando la pared interna y externa de los pilares externo e interno. Ambas filas están distantes en sus bases, uniéndose por arriba, lo que hace que el túnel tenga una cierta forma piramidal.
Están situadas en la parte media del órgano de Corti y son de forma piramidal y son un tanto particulares o propias de esta zona. Los dos pilares, externo e interno, formados por estas cc, delimitan junto con la membrana basilar el túnel de Corti. Están situadas en la parte media del órgano de Corti y son de forma piramidal. Están dispuestas en dos filas, externa e interna, formando la pared interna y externa de los pilares externo e interno. Ambas filas están distantes en sus bases, uniéndose por arriba, lo que hace que el túnel tenga una cierta forma piramidal.
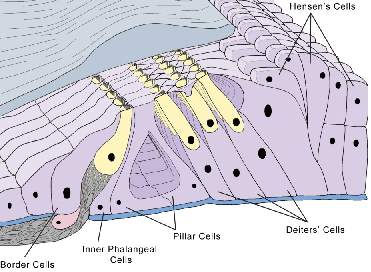
Células de Deiters, son las cc de sostén para las CCE. Están situadas por fuera de los pilares. Tienen forma fusiforme, delimitando entre ellas los espacios de Nuel. Los polos superiores de estas cc forman entre ellos unos espacios libres, en forma de cúpula abierta hacia arriba, para recibir a las cc ciliadas. En su cima son afiladas, entrando a formar parte de la membrana reticular.
Las cc de sostén de las CCI, están situadas por dentro de los pilares y son análogas las cc de Deiters.
 - Células sensoriales.
- Células sensoriales.
Son cc ciliadas, asientan sobre la membrana basilar, siguen una disposición espiral a lo largo de la cóclea y están dispuestas en dos grupos a cada lado de los pilares: células ciliadas internas y células ciliadas externas. Ambas poblaciones celulares son funcional e histológicamente diferentes.
Poseen un cuerpo globuloso, con estereocilios en su polo superior ordenados linealmente sin hacer contacto con la superficie inferior de la membrana tectoria.
Las CCI se calcula que pueden ser unas 3.500 y están colocadas en una sola fila. Son las cc sensitivas auditivas verdaderas y proporcionan la transducción mecánico-eléctrica, formando sinapsis con el 95% de los axones que constituyen la vía aferente auditiva que va a parar al cerebro.
Las CCE son más numerosas, unas 13.000-20.000, y forman tres filas en el hombre. En algunos animales están dispuestas en más filas. Están dispuestas en forma alineada, casi en línea recta, y están apoyadas en las cc de Deiters. El extremo inferior de las cc ciliadas es redondeado y reposa sobre las cúpulas receptoras que forman las cc de Deiters. El extremo superior de las cc ciliadas está encastrado en la membrana reticular.
Estas cc ciliadas, como las de los neuroepitelio del vestíbulo y CS, poseen dos tipos de cilios: unos cortos y numerosos, llamados esterocilios y otro más largo y único, llamado kinocilio, que en el caso del Órgano de Corti existe sólo en forma rudimentaria.
Cada CCI, tienen forma piriforme, posee en su superficie múltiples esterocilios ordenados en hilera, alrededor de unos 60-100 por cc. Las oscilaciones de los esterocilios provocaran la activación celular, y, por tanto, el envío del mensaje auditivo al encéfalo. En su porción basal hacen sinapsis directamente con las fibras aferentes que se proyectan al SNC. Las fibras nerviosas y las cc de sostén rodean la superficie basolateral de forma que la CCI no llega a hacer contacto con la membrana basilar. Estas cc poseen uniones estrechas (tight junctions) con las cc de sostén a nivel de la lámina reticular. Su superficie apical está separada de las otras CCI por cc de sostén. Sin embargo, las CCI contactan con sus vecinas en las zonas en donde el cuerpo se ensancha a nivel del núcleo. Por lo tanto cabría la posibilidad de una comunicación directa entre ellas a este nivel que aún no ha podido ser demostrada.
 Las CCE poseen entre 100 y 120 cilios por cc. Los esterocilios se ordenan en V y están firmemente anclados a la membrana tectoria. Estas cc son alargadas y cilíndricas con el núcleo en situación más basal. Tienen menor diámetro que las CCI, su superficie apical contacta con las cc de sostén mediante uniones estrechas (tight junctions) y uniones adherentes (adherens junctions). Cada CCE está envuelta en su base por una cc de Deiters, algunas de estas cc presentan una prolongación alargada que asciende formando parte de la lámina reticular. A diferencia de las CCI, las CCE tienen libres sus superficies laterales que sólo contactan con el fluido circundante. Esta característica imposibilita la comunicación directa entre ellas pero, no obstante, facilita su función contráctil. Cave recordar, que si bien ambos tipos de cc sensoriales tiene una función de traductor mecano-eléctrico del sistema auditivo, las CCE actúan además con transductores electromecánicos, es decir como cc motoras. Estas CCE pueden estar acopladas a cc de sostén a través de uniones de hendidura (gap junc). Los cilios de las CCE se disponen en V abierta hacia la columela, disposición que es aún más pronunciada en la base del caracol donde se perciben las frecuencias altas. Más cerca de la punta, la forma en V es más pequeña y cerrada.
Las CCE poseen entre 100 y 120 cilios por cc. Los esterocilios se ordenan en V y están firmemente anclados a la membrana tectoria. Estas cc son alargadas y cilíndricas con el núcleo en situación más basal. Tienen menor diámetro que las CCI, su superficie apical contacta con las cc de sostén mediante uniones estrechas (tight junctions) y uniones adherentes (adherens junctions). Cada CCE está envuelta en su base por una cc de Deiters, algunas de estas cc presentan una prolongación alargada que asciende formando parte de la lámina reticular. A diferencia de las CCI, las CCE tienen libres sus superficies laterales que sólo contactan con el fluido circundante. Esta característica imposibilita la comunicación directa entre ellas pero, no obstante, facilita su función contráctil. Cave recordar, que si bien ambos tipos de cc sensoriales tiene una función de traductor mecano-eléctrico del sistema auditivo, las CCE actúan además con transductores electromecánicos, es decir como cc motoras. Estas CCE pueden estar acopladas a cc de sostén a través de uniones de hendidura (gap junc). Los cilios de las CCE se disponen en V abierta hacia la columela, disposición que es aún más pronunciada en la base del caracol donde se perciben las frecuencias altas. Más cerca de la punta, la forma en V es más pequeña y cerrada.
Otra característica de los cilios de ambos tipos de cc ciliadas es que los cilios que posee cada cc están unidos entre sí por puentes glicoproteícos que permiten que todos los elementos de una misma cc se desplacen conjuntamente.
 - Otras células del órgano de Corti.
- Otras células del órgano de Corti.

▪ Membranas.
- La membrana de Reissner, separa el canal coclear de la rampa vestibular, constituye el techo del canal coclear.
En su cara inferior, coclear o endolinfática, está recubierta por grandes cc poligonales, aplanadas, monoestratificadas que están conectadas entres sí por zónulas de oclusión y zónulas de adherencia. Poseen microvellosidades. El citoplasma de estas cc es relativamente pobre en orgánulos celulares y este dato, así como la falta de irrigación vascular directa, indica que la membrana de Reissner no desempeña más que un papel secundario en la regulación activa de la composición de la endolinfa.
En su cara superior, vestibular, o perilinfática, se compone de cc muy planas, endoteliformes que no forman un conjunto compacto siendo una continuación de las que revisten la rampa o el espacio vestibular. El resto de la cavidad perilinfática está igualmente recubierta de cc similares.
- Membrana reticular. Es una sustancia cementante homogénea, formada por fibrillas de colágeno. Esta membrana recorre todo el órgano espiral en forma de red y por sus orificios pasan los cilios de las cc ciliadas, además une los polos apicales de las cc de Deiters y de las CCE.
- Membrana tectoria. Es una membrana espesa, compuesta de una sustancia amorfa, gelatinosa, recorrida por minúsculas fibrillas, con el mismo peso específico que la endolinfa. Está sólidamente fija al limbo de la lámina espiral. Recubre por fuera el órgano de Corti y juega un papel muy importante en la fisiología de la audición. Los cilios de la cc sensoriales penetran en esta membrana.
 ▪ Histología de la estría vascular.
▪ Histología de la estría vascular.
Es la estructura responsable de la producción de la endolinfa. Está colocada sobre el ligamento espiral, tapizando los dos tercios superiores de la pared externa del canal coclear.
Histológicamente está formada por un epitelio estratificado con abundantes capilares en su interior. En su epitelio se pueden considerar tres tipos de cc: basales, intermedias y superficiales.
Células basales. Tienen forma poligonal y se sustentan sobre una membrana basal mal definida que a veces sólo se evidencia como un depósito de material amorfo entre las cc basales y los fibroblastos del tejido conjuntivo que forma el ligamento espiral. Estas cc muestras en su superficie apical una depresión en forma de cáliz, en la que se alojan las cc intermedias y las superficiales. Están unidas por desmosomas, tienen un núcleo voluminoso y un número escaso de organelos. Están unidas a las cc basales, a las cc intermedias y a los fibroblastos del ligamento espiral por medio de uniones gap. Se interpretan como cc madre.
Células intermedias. Tienen forma estrellada y están ubicadas entre las basales y las superficiales. Rodean a los capilares. Su núcleo es central y de forma esférica, su citoplasma es claro. Embriológicamente se cree que son melanocitos que llegan a la estría procedentes de cc emigradas desde la cresta neural. Contienen una importante dotación enzimática que les permite obtener energía de sustratos alternativos como los lípidos y actuar ejerciendo una acción desoxificadora de los restos de la oxidación. Al tener la capacidad de captar radicales libres, se ha sugerido que estas cc protegerían a la estría vascular del estrés y suministrarían energía alternativa en caso de reducción del aporte sanguíneo.
Células marginales. También denominadas superficiales tienen su polo apical liso, con escasos microvilli y su polo basal irregular que se acopla en la concavidad formada por el cáliz de la cc basal caracterizado por tener invaginaciones profundas. El polo apical muestra una morfología penta o hexagonal y es ligeramente convexo. En sus extremos apicales están unidas entre ellas por complejos de unión. Su núcleo voluminoso es de cromatina laxa y de morfología esférica u ovoidea. Su citoplasma contiene vesículas de micropicnocitosis, cisternas de retículo endoplásmico rugoso, lisosomas, cuerpos multivesiculares, un aparato de Golgi poco desarrollado y algunos enclaves lipídicos y gránulos de lipofucsina. Las mitocondrias se sitúan rodeando al núcleo pero se localizan preferentemente en las prolongaciones distales que dan origen a las invaginaciones del polo basal. Estas cc son las responsables del transporte de K a la endolinfa.
Para continuar con este capítulo descargar tema completo![]()