-
Embriología de la glándula tiroides.
-
Embriología de las glándulas paratiroides.
 EMBRIOLOGÍA DE LA GLÁNDULA TIROIDES.
EMBRIOLOGÍA DE LA GLÁNDULA TIROIDES.
Los dos órganos de secreción interna más precoces en esbozarse en el embrión humano son el tiroides y la adenohipófisis.
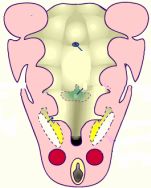
En el embrión humano de 13-20 somitos, horizontales XI-XII de Streeter, al final de la tercera semana, se observa ya un primordio de la glándula tiroides de origen endodérmico formado por una invaginación impar de epitelio endoblástico, situado en el suelo faríngeo debajo de la lengua, es decir en la línea media del suelo de la boca primitiva, en el punto donde se situará el foramen cecum. A la altura de esta depresión las cc del epitelio faríngeo son más altas que en las regiones vecinas. Su situación embriológica exacta corresponde a la parte medioventral del área mesobranquial, en la unión de los arcos viscerales II y III, región denominada cúpula, que se localiza caudal al área deltoide, o de confluencia de los arcos I y II. La porción rostral del esbozo tiroideo forma una prominencia progresivamente mayor en el suelo de la boca primitiva por el incremento del mesénquima subyacente.
Este primordio, formado por una evaginación endodérmica, se va deprimiendo en el mesénquima subyacente mesobranquial en sentido caudal. Se sitúa en inmediata vecindad del saco aórtico junto al primordio que va a formar el corazón y como éste experimenta un desplazamiento caudal alargándose. Al irse alargando, forma en su extremo inferior un divertículo endodérmico que al mes se va a bilobular delante de la cuarta bolsa faríngea.
 En su emigración el primordio descendente va dejando un tallo que lo une al suelo faríngeo, que se denomina conducto tirogloso de Hiss. Este tallo o conducto en el 2º mes se fragmenta y se disuelve, dejando en su punto de origen, que corresponde el dorso lingual, un hoyuelo denominado foramen caecum. El conducto puede, a veces, no reabsorberse, persistiendo en su totalidad, o dejando restos celulares que pueden diferenciarse en tejido tiroideo, originando de esta forma tiroides accesorios. Parte del extremo caudal del conducto suele persistir, dando origen a una parte de la glándula denominada lobus piramidalis o pirámide.
En su emigración el primordio descendente va dejando un tallo que lo une al suelo faríngeo, que se denomina conducto tirogloso de Hiss. Este tallo o conducto en el 2º mes se fragmenta y se disuelve, dejando en su punto de origen, que corresponde el dorso lingual, un hoyuelo denominado foramen caecum. El conducto puede, a veces, no reabsorberse, persistiendo en su totalidad, o dejando restos celulares que pueden diferenciarse en tejido tiroideo, originando de esta forma tiroides accesorios. Parte del extremo caudal del conducto suele persistir, dando origen a una parte de la glándula denominada lobus piramidalis o pirámide.
 El divertículo final del conducto, que origina el tiroides, se denomina vesícula tiroidea. Esta comienza enseguida a tener una activa proliferación celular ordenada en un principio mediante agrupamientos cordonales celulares, intercalados de conectivo vascular. Posteriormente puede apreciarse que la disposición celular es en forma de unas cavidades a cuyo alrededor se disponen las cc, siendo esta disposición el inicio o esbozo de los folículos. Posteriormente, unas dos semanas después, los cordones celulares irán tomando forma de auténticos folículos repletos de líquido coloide, el tiroides ha aumentado de taño y es ya captador de I. Se considera que la glándula comienza a ser funcional en el feto de tres meses, si bien en la 9ª semana ya es capaz de sintetizar hormonas yodadas. Bajo una estimulación intensa, el tiroides del adulto puede formar más folículos nuevos.
El divertículo final del conducto, que origina el tiroides, se denomina vesícula tiroidea. Esta comienza enseguida a tener una activa proliferación celular ordenada en un principio mediante agrupamientos cordonales celulares, intercalados de conectivo vascular. Posteriormente puede apreciarse que la disposición celular es en forma de unas cavidades a cuyo alrededor se disponen las cc, siendo esta disposición el inicio o esbozo de los folículos. Posteriormente, unas dos semanas después, los cordones celulares irán tomando forma de auténticos folículos repletos de líquido coloide, el tiroides ha aumentado de taño y es ya captador de I. Se considera que la glándula comienza a ser funcional en el feto de tres meses, si bien en la 9ª semana ya es capaz de sintetizar hormonas yodadas. Bajo una estimulación intensa, el tiroides del adulto puede formar más folículos nuevos.
 Hacia la 7ª semana la vesícula tiroidea alcanza su localización pretraqueal definitiva, aún cuando su vecino, el esbozo cardíaco, continúa descendiendo. Con el desarrollo del cuello embrionario, el corazón y los grandes vasos se separan del tiroides, pero algunos pequeños islotes de tejido tiroideo pueden permanecer adheridos a la aorta o a sus ramas, lo que explica la ocasional presencia de tejido tiroideo en la cavidad torácica.
Hacia la 7ª semana la vesícula tiroidea alcanza su localización pretraqueal definitiva, aún cuando su vecino, el esbozo cardíaco, continúa descendiendo. Con el desarrollo del cuello embrionario, el corazón y los grandes vasos se separan del tiroides, pero algunos pequeños islotes de tejido tiroideo pueden permanecer adheridos a la aorta o a sus ramas, lo que explica la ocasional presencia de tejido tiroideo en la cavidad torácica.
El desarrollo de los folículos, el coloide y el desarrollo anatómico de la glándula en general es estimulado por la TSH fetal, pues la placenta es impermeable a la TSH, T3 y T4 maternas. La TSH comienza a segregarse en el feto a la 10ª semana. Antes de la 10ª semana puede detectarse en el feto TBG y entre la 10ª y 11ª semana comienza la síntesis de T4. El metabolismo productor-destructor de T4 intrafetal es distinto al del adulto con unos niveles marcadamente superiores. Aunque la cantidad de hormonas tiroideas secretadas por el feto es desconocida, se sabe que es suficiente, ya que animales completamente tirectomizados pueden dar a luz recién nacidos totalmente normales. Al final de embarazo la aportación de hormonas tiroideas maternas es prácticamente nula y, por el contrario, es posible que las hormonas fetales incrementen el contenido hormonal de la madre.
Las hormonas tiroideas condicionan la última fase de la maduración esquelética, pulmonar, cerebral, función intelectual y organización armónica del sistema nervioso (cretinismo), no obstante, la observación de neonatos con ausencia de función tiroidea revela que el desarrollo somático intrafetal es independiente de la hormona.
EMBRIOLOGÍA PARATIROIDES.
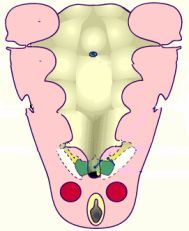
Las paratiroides inferiores derivan de la porción dorsal del endodermo de la 3ª bolsa branquial, y las paratiroides superiores de la porción dorsal del endodermo de la 4ª bolsa branquial junto a los lóbulos laterales del tiroides. El tejido paratiroideo comienza a desarrollarse cuando el embrión mide de 8 a 10mm, tiene entonces 5 semanas.
Empiezan a desarrollarse en la 5ª semana de vida. En embriones humanos de 7 mm se observan unas expansiones epiteliales bilaterales, o zonas paratidogénicas, en el endodermo dorsal de las bolsas faríngeas III y IV. La III bolsa dará origen a las paratiroides inferiores y la IV bolsa a las superiores.
En embriones de 13 mm en la III bolsa faríngea se distingue una porción craneal, otra caudal y otra intermedia, siendo en la porción craneal donde se observan los esbozos partiroideos. Los esbozos paratiroideos forman cordones epiteliales sólidos denominados cropúsculos epiteliales.
El complejo paratiroides inferiores realizan un descenso común con el timo, se despegan de la pared faríngea y progresan hacia la región caudal y mediana. En el embrión de 20mm, la regresión cefálica del timo genera su separación de las paratiroides inferiores, las cuales quedan en la región anterior o posterolateral de los polos inferiores de los lóbulos tiroideos o a nivel de los ligamentos tirotímicos. Esta forma de migración hace que su localización definitiva sea muy variable pudiendo localizarse desde el ángulo de la mandíbula hasta el mediastino anterior. Por tanto las paratiroides inferiores tienen una localización más inconstante que las superiores debido a que realizan una migración caudal más rápida y larga junto con el tiroides inferior y el timo. En la mayoría de los tiroides se localizan inferior, lateral y posterior mente al polo inferior de lóbulo tiroideo, por debajo del nivel de la arteria tiroidea inferior.
Las paratiroides superiores siguen la migración tiroidea que se dirige hacia la parte lateral de de los esbozos tiroideos medianos principales; el descenso es corto. Se encuentran situadas junto a los cuerpos ultimofaríngeo, cuerpo ultimobranquial o glándula ultimobranquial y ambas estructuras descienden de forma conjunta y se separan al incorporarse los cuerpos últimobranquiales a la glándula tiroides. Debido a ello, en el adulto, la paratiroides superior mantiene una íntima relación con el tubérculo de Zuckerkandl. Al tener un recorrido embriológico más corto que las inferiores, su variabilidad en localización es menor y su ubicación topográfica es más estable. Quedan en contacto con la cara posterior de los lóbulos tiroideos a nivel de su tercio medio, localizándose lo más frecuentemente a 1 cm por encima de la intersección entre el nervio laríngeo recurrente y la arteria tiroidea inferior, cerca de la articulación crirco-tiroidea. Las paratiroides superiores son cruzadas por las inferiores tras el descenso paratímico.
En embriones de 15 mm. histológicamente las cc tiene ya el aspecto de cc paratiroideas y se pueden apreciar las paratiroides ya totalmente aisladas de la faringe y ocupando sus posiciones definitivas. Aunque las bolsas faríngeas se separan de la faringe, las glándulas quedan unidas a la faringe mediante un conducto faringobranquial, más estrecho en la bolsa III que en la IV.