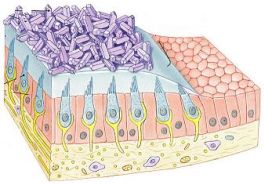
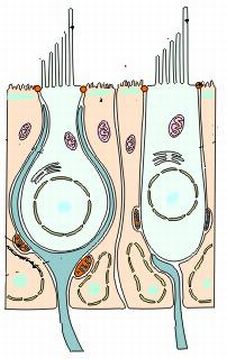
Las paredes del laberinto membranoso están formadas por tres capas histológicas. Una membrana basal intermedia recubierta en su cara interna y externa por epitelio.
-
La membrana basal o cubierta conjuntiva está formada por fibras conjuntivas y elásticas.
-
La cara interna de la membrana basal está tapizada de un epitelio pavimentoso simple. Este epitelio en determinadas zonas experimenta un engrosamiento formado por cc de tipo diferenciado que son las cc sensoriales: son las zonas en las que se ubican los receptores neurosensoriales del laberinto posterior. Estos receptores son las maculas del utrículo y el sáculo y las crestas ampulares de los canales semicirculares. Entre el epitelio sensorial de los receptores periféricos y el epitelio pavimentoso que reviste el interior de las cavidades, existe una zona intermedia de transición, constituida por un epitelio de aspecto secretor denominado plano semilunatum, muy vascularizado y que es productor de endolinfa.
-
La cara externa de la membrana basal está recubierta de un endotelio de cc perilinfáticas.
La capa externa y la basal, en las zonas donde asientan los receptores están perforadas por las terminaciones nerviosas dendríticas de los nervios vestibulares.
 MÁCULAS.
MÁCULAS. Constituyen los receptores neurosensoriales del utrículo y el sáculo, hay dos máculas utriculares y otras dos máculas saculares (una en cada lado). En el capítulo anterior se ha descrito su estructura macroscópica.
Microscópicamente están constituidas por un epitelio basal formado por cc cúbicas de sostén que descansan sobre la membrana basal y por encima de estas cc se disponen las cc sensoriales ciliadas. Las cc de sostén fijan las cc sensoriales a la membrana basal.
La membrana basal está situada por encima de tejido conjuntivo y tiene una serie de agujeros por donde penetran las fibras nerviosas con destino a las cc ciliadas neurosensoriales.
Por encima de las cc sensoriales, reposando sobre el epitelio sensorial, se dispone una masa semigelatinosa denominada membrana otolítica, estatoconial o estatolítica. Tiene un espesor de cincuenta micrómetros y alrededor de un milímetro cuadrado de superficie. En su base presenta orificios para el paso de los cilios de las cc sensoriales que penetran en ella.
La masa de la membrana tiene como componentes más importantes glicoproteínas y sulfomucopolisacáridos. Estructuralmente está formada por una densa red de filamentos arrosariados y columnares que terminan anclándose alrededor de la la base de las microvellosidades de las cc de sostén y ocasionalmente en las cc sensoriales. Este tipo de estructura facilita la fijación de la membrana otolítica al epitelio basal.
Esta masa está tapizada por una gran cantidad de pequeños cristales de carbonato cálcico y proteínas denominados otolitos, otoconias u estatoconias incrustados en la misma. La membrana otolítica, al tener incrustados los otolitos, es tres veces más densa y viscosa que la endolinfa.
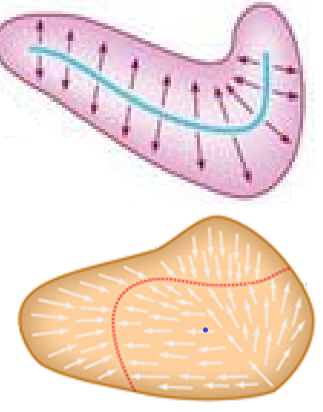 A lo largo del centro de cada membrana otolítica se extiende una línea curva virtual, denominada estríola, que la diferencia en dos partes y que es el límite morfológico y funcional de la detección de aceleración en uno u otro sentido, ya que los cilidos de las cc ciliadas están orientadas en direccion opuesta a cada lado de la estriola, como explicaremos al tratar la fisiología del sistema otolítico.
A lo largo del centro de cada membrana otolítica se extiende una línea curva virtual, denominada estríola, que la diferencia en dos partes y que es el límite morfológico y funcional de la detección de aceleración en uno u otro sentido, ya que los cilidos de las cc ciliadas están orientadas en direccion opuesta a cada lado de la estriola, como explicaremos al tratar la fisiología del sistema otolítico.
Constituyen el órgano receptor de los canales semicirculares y está alojado en la pared interna de cada una de las ampollas.
En el interior de cada ampolla el tejido conjuntivo de la pared interna forma un relieve o protuberancia trasversal en forma de media luna que será la base de la cresta acústica o cresta ampular. Forma un ángulo recto con la pared de la misma, es decir, su orientación es perpendicular al conducto. Ocupa como una tercera parte de luz de la ampolla. Histológicamente, de la misma forma descrita en la máculas. está formada por una membrana basal atravesada por fibras nerviosas sobre la que se asientan cc de sostén y cc ciliadas sensoriales ciliadas distribuidas regularmente en forma de mosaico. Por debajo de la membrana basal se sitúan los nervios y los vasos dentro del tejido conectivo de soporte.
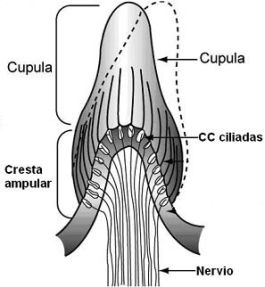 Sobre el neuroepitelio de cada cresta se encuentra una formación homogénea, de consistencia gelatinosa con una estructura similar a la membrana otolítica de utrículo y sáculo, pero con forma de cúpula. A esta masa se la denomina cúpula ampular y en ella se enclavan los cilios de las cc sensoriales. Los cilios se introducen por unos conductillos verticales en la masa gelatinosa de la cúpula.
Sobre el neuroepitelio de cada cresta se encuentra una formación homogénea, de consistencia gelatinosa con una estructura similar a la membrana otolítica de utrículo y sáculo, pero con forma de cúpula. A esta masa se la denomina cúpula ampular y en ella se enclavan los cilios de las cc sensoriales. Los cilios se introducen por unos conductillos verticales en la masa gelatinosa de la cúpula. La cúpula tiene la misma densidad que la endolinfa y puede moverse, pues a diferencia de la membrana otolítica ésta no tiene otolitos. La cúpula es una sustancia gelatinosa en cuya composición figuran un 90% de proteínas tipo mucopolisacáridos y un armazón de fibras colágenas dispuestas en todas las direcciones a forma de red. Según esto, hay una gran diferencia entre las máculas y las cúpulas, las máculas tienen una densidad superior a la del líquido endolinfático, mientras que la densidad de las cúpulas es similar a la de éste. Esta diferencia, como hemos dicho, se debe a que las cúpulas no tienen otolitos.
 En la periferia del epitelio sensorial la histología de la cresta cambia a un epitelio de transición entre el sensorial y el resto del epitelio de recubrimiento de la ampollas. Esta zona se denomina planum semilunatum. Tiene forma de media luna y su epitelio se caracteriza por que sus cc tienen características secretoras, van disminuyendo de altura a medida que se alejan del epitelio sensorial, tienen menos microvelli que las cc de soporte, su núcleo es oval situado cerca de la membrana basal. Su membrana plasmática inferior presenta numerosos pliegues involucrados en el metabolismo de agua e iónico, como también sucede con las cc oscuras que describiremos más abajo.
En la periferia del epitelio sensorial la histología de la cresta cambia a un epitelio de transición entre el sensorial y el resto del epitelio de recubrimiento de la ampollas. Esta zona se denomina planum semilunatum. Tiene forma de media luna y su epitelio se caracteriza por que sus cc tienen características secretoras, van disminuyendo de altura a medida que se alejan del epitelio sensorial, tienen menos microvelli que las cc de soporte, su núcleo es oval situado cerca de la membrana basal. Su membrana plasmática inferior presenta numerosos pliegues involucrados en el metabolismo de agua e iónico, como también sucede con las cc oscuras que describiremos más abajo.
Es una estructura gelatinosa que se coloca a modo de casco por encima del epitelio sensorial de la cresta. Se extiende desde la superficie del epitelio sensorial hasta el techo y las paredes laterales de la ampolla cerrando hermeticamente la luz de la ampolla, de esta forma divide la luz de cada canal en dos compartimentos independientes a dada lado de la ampolla, uno de ellos próximo al utrículo y el otro más alejado.
Está compuesta por filamentos de unos 30-40 A de espesor, dentro de los cuales existe un sistema de canales que los recorren en toda su extensión, desde la base al vértice y bordes laterales, y dentro de ellos se introducen los haces de los cilios sensoriales.
Se han encontrado en su composición sulfomucopolisacáridos secretados por las cc de soporte y glusaminoglicanos que son son polímeros no ramificados de azúcares.
Entre la superficie cóncava de la cúpula y la convexa correspondiente al epitelio sensorial de la cresta existe un pequeño espacio subcupular de unas 4 micras de espesor. Los cilios de las cc sensoriales atraviesan este espacio para penetrar en una serie de canalículos preformados en el seno de la sustancia cupular atravesándola desde su superficie inferior hasta su vértice. Estos canalículos van disminuyendo su calibre según ascienden y algunos no son del todo rectilíneos pues tienden a converger hacia el vértice.
CÉLULAS SENSORIALES DE LOS RECEPTORES VESTIBULARES.
Las cc sensoriales de los receptores vestibulares situados en las crestas ampullares y en las máculas del utrículo y sáculo, son cc ciliadas. Se comenzaron a conocer detalladamente en la década de los 50 con la llegada de la microscopia electrónica. Hoy se conoce ya muy bien su estructura.
La actividad, morfología y polarización morfofuncional de las cc ciliadas de las máculas son similares a las de las cc ciliadas de las crestas.
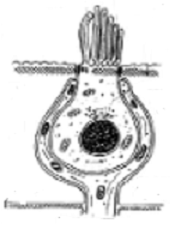
Son cc con una membrana celular de 70 À. En la cc sensorial se pueden diferenciar diferentes zoas.
En su parte superior presenta una formación cuticular y los cilios. Esta es una primera zona, o zona cuticular superficial. Los cilios se encuentran en el polo apical de estas cc. y se introducen en la sustancia gelatinosa que recubre la superficie celular, estando en contacto con la endolinfa.
Debajo de esta zona se encuentra la denominada zona intermedia caracterizada por la presencia de organitos endocelulares y otras formaciones de importancia funcional celular.
Le sigue en profundidad la tercera zona o zona perinuclear que rodea al núcleo, caracterizada por ser pobre en mitocondrias.
Finalmente, la más profunda es la zona neurosensorial, donde la cc toma contacto con la terminación nerviosa que le corresponde pero sin que ésta penetre en el citoplasma.
Los cilios son de dos tipos:
- Un solo kinocilio (o cinetocilio) por cc, que es el cilio activo de la misma. Tiene un corpúsculo basal y la estructura interna típica de un cilio formada por microtúbulos que discurren longitudinalmente a lo largo del cilio.
- Múltiples esterocilios inactivos por cada cc que son prolongaciones celulares de menor grosor y longitud que el kinocilio implantadas en la zona apical cuticular. Son de longitud desigual, estando los más largos más cerca del kinocilio y va disminuyendo su longitud según se alejan del kinocilio. Son estructuras relativamente rígidas que pivotan sobre su base cuando son empujados externamente.
La situación del kinocilio con relación a los esterocilios en cada cc sensorial es fundamental para entender la polaridad celular. El kinociio constituye el sustrato morfofuncional para la existencia de una determinada polaridad celular, lo que explica las leyes I y II de Edwald (ver en capítulo 05.1ª.05); la polaridad de la cc sensorial en cada momento depende de fenómenos micromecánicos que mas abajo explicamos. El kinocilio no tiene plano cuticular, su pie basal está orientado en dirección opuesta al los esterocilios y carece de movilidad.
En el utrículo, los kinocilios de las cc situadas a cada uno de los dos lados de la línea de la estriola se muestran los un lado en frente de los del otro lado. Esta línea comienza en la extremidad anterior de la mácula y termina en su borde medial, siguiendo un trayecto curvilineo. En el sáculo están colocados alejados de la línea de la estriola, teniendo en cuenta que la estriola aquí comienza en la parte más postero-superior de la mácula desde donde toma un trayecto en S itálica y se dirige hacia abajo, aproximándose primero al plano anterior y después al posterior.
En las crestas ampulares el kinocilio se sitúa en el CSH más próximo al utrículo, y en los CS verticales, en el lado opuesto, está más alejado del utrículo.
 Las cc sensoriales del laberinto posterior son mecanorreceptores especializados en la captación de desplazamientos inerciales de masas para transformarlos en impulsos nerviosos. El estímulo adecuado es idéntico para todas ellas y está constituido por fuerzas que producen la flexión de los cilios. Ahora bien, las fuerzas estimulatorias son diferentes para las crestas y para las máculas. En las crestas son aceleraciones angulares y en las máculas son aceleraciones lineales y la fuerza de la gravedad.
Las cc sensoriales del laberinto posterior son mecanorreceptores especializados en la captación de desplazamientos inerciales de masas para transformarlos en impulsos nerviosos. El estímulo adecuado es idéntico para todas ellas y está constituido por fuerzas que producen la flexión de los cilios. Ahora bien, las fuerzas estimulatorias son diferentes para las crestas y para las máculas. En las crestas son aceleraciones angulares y en las máculas son aceleraciones lineales y la fuerza de la gravedad. Las cc sensoriales tienen una actividad en reposo de aproximadamente 90 impulsos por segundo. Esta actividad es modulada, aumentado o disminuyendo al producirse un estímulo.
Cada esterocilio posee nos 3.000 filamentos de actina que disminuyen en grosor a medida que se aproximan a la superficie apical de la cc y por abajo. De los 3.000 solamente unos 20 llegan a alcanzar desde su raíz el plano cuticular. Además de la actina posee otras proteínas contráctiles como la fibrina y la tropomiosina con una polaridad uniforme dirigida hacia el plano cuticular.
Cada cc sensoriales posee unos 60 a 100 esterocilios insertados en el plano cuticular de la superficie apical celular que se disponen en 5 hileras regulares de 8 cilios cada una. Su longitud se incrementa escalonadamente de tal forma que la hilera más alta se sitúa en la extremidad de la cc más próxima al kinocilio. Son más cortos en las máculas utriculosaculares (12 micromicras) que en las cretas ampulares (30 micromicras). En cada cilio se puede considerar una porción intracelular y otra mayor extracelular, continuación una de la otra. En el interior del cilio se puede apreciar una fina estructura central o fibrilla central y una porción periférica a modo de vaina de aquella. Por tanto cada cc posee por término medio unos 40 esterocilios y un kinocilio.
El microscopio electrónico permite distinguir dos tipos de cc sensoriales (Wersall):
 - Células de tipo I.
- Células de tipo I. Su cuerpo tiene forma piriforme-globulosa, o forma de copa, en el que se distingue un extremo inferior redondeado, un cuerpo que se va adelgazando de abajo hacia arriba y una cabeza ancha y aplanada de la cual arranca el haz ciliar. En el polo superior de la cc están los cilios y en su base, con forma de cáliz, está en gran parte rodeada de una terminación nerviosa aferente. Tienen una sola terminación nerviosa por cc, si bien en algunas cc de las máculas, una terminación nerviosa puede ser común para 3 ó 4 cc. Este tipo de cc se localizan por todo el epitelio sensorial, pero son más abundantes en la cúspide de la cresta ampular y en el centro de las máculas.
En el extremo apical o cabeza, cono hemos dicho, hay un plano cuticular. El plano cuticular ocupa una gran parte de la superficie celular y en él se encuentra una red filamentosa, gránulos densos y una serie de proteínas motoras como actina, miosina y tubulina.
El haz ciliar está formado por 60-100 cilios que se insertan en el plano cuticular Están dispuestos en hileras regulares y su longitud se incrementa escalonadamente de tal forma que la hilera más alta se sitúa en el extremo de la cc más próximo al kinocilio. En general son más estrechos en su base que en el vértice. Los esterocílios de las cc de tipo I son más numerosos y más gruesos que los de las cc de tipo II, lo que hace que las cc de tipo I sean un receptor más dinámico, con respuestas más rápidas que las de tipo II.
Cada esterocilio está recubierto de un glucocáliz de carbohidratos que tiene las siguientes funciones:
- Establecer entre los esteroilios una carga electrostáticamente negativa de repulsión.
- Mantener mediante una serie de interconexiones una armonía y simetría de todo el haz de esterocílios.
- Hacer que el glucocáliz del esterocilio mantenga una disposición de unidad morfofuncional con los componentes filamentosos de la membrana otológica, de la tectoria y de la cúpula de los CS.
El conjunto de proteínas que constituyen el esterocilio y el plano cuticular actúan como un sistema de palancas que transmite los estímulos mecánicos que recibe. Hemos de señalar que los esterocilios vestibulares tienen menor rigidez en el momento de su inflexión que los cocleares.
El núcleo de estas cc es grande, esférico y está situado en el centro de la cc. Su citoplasma muestra una sustancia homogénea con abundantes mitocrondrias.
En el polo basal de estas cc se sitúa el área sináptica; la terminación nerviosa eferente tiene forma abotonada, con numerosas vesículas sinápticas y una hendidura sináptica de unos 200 A. Como ya hemos dicho el cáliz neural aferente suele rodear a una o dos cc sensoriales envolviéndolas hasta la proximidad del cuello.
 - Células de tipo II.
- Células de tipo II. Filogenéticamente son cc más antiguas que las de tipo I.
Predominan en las zonas periféricas de la cresta ampular e igualmente en la periferia de las máculas.
Tienen forma cilíndrica. La disposición del kinocilio, esterocilio y de los restantes elementos celulares es la misma que en las de tipo I, diferenciándose exclusivamente en su inervación. Su base está en relación con múltiples y pequeñas terminaciones nerviosas de forma abotonada, aferentes y eferentes situadas en el tercio inferior de la cc sensorial. Las eferentes no contactan con la cc, sino que lo hacen con la fibra aferente. Las terminaciones aferentes carecen de vesículas sinápticas, que por otra parte son muy abundantes en las eferentes.
La diferencia en los terminales periféricos de los axones sensoriales vestibulares de cada uno de los dos tipos de cc hace que las células tipo I responden a una estimulación vestibular más intensa, mientras que las células tipo II responden a estímulos más débiles. Sin embargo, los mecanismos de respuesta son similares para ambos tipos de cc.
En el epitelio sensorial de las máculas se han descrito un tercer tipo de cc que se han denominado otolitocitos, considerándolas productoras de otolitos.
Las cc de sostén o de soporte se encuentran situadas entre la membrana basal y la superficie libre del epitelio. Están rodeadas por cc sensoriales y terminaciones nerviosas.
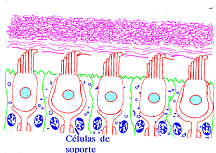 Tienen forma prismática o cilíndrica irregular en su superficie por amoldarse a la forma de las vecinas. En su superficie están unidas en su parte apical a las cc sensoriales mediante complejos de unión y desmosomas.
Tienen forma prismática o cilíndrica irregular en su superficie por amoldarse a la forma de las vecinas. En su superficie están unidas en su parte apical a las cc sensoriales mediante complejos de unión y desmosomas. Su superficie apical presenta un plano cuticular con amplias soluciones de continuidad en el centro donde puede implantarse un corpúsculo basal del cual arranca un kinocilio rudimentario.
La estructura citoplásmica de estas cc es muy compleja, rica en mitocondrial, complejos de Golgi, lisosomas y ribosomas, por lo que se les ha atribuido una función secretora y metabólica al servicio de las cc sensoriales y de sus prolongaciones dendríticas.
El núcleo es oval y está situado en el extremo inferior de la cc, situándose por debajo de la altura a la que están situados los núcleos de las cc sensoriales.
UNIDADES DE CÉLULAS OSCURAS.
En la periferia de los receptores sensoriales de la mácula del utrículo y de las crestas ampulares, en el plano semilunatum, existen las denominadas unidades de cc oscuras. Reciben este nombre por la mayor densidad electrónica de su citoplasma. Estas unidades es situadas lateralmente a las cc transicionales, que las separan de la zona en la que se ubican las cc receptoras y las cc de soporte. Las unidades de cc oscuras son las responsables de mantener la homeostasis de la endolinfa.
Histológicamente están constituidas por un epitelio formado por las cc oscuras, bajo el cual se dispone un tejido conjuntivo que contiene cc pigmentadas más o menos relacionadas con las cc oscuras. En las crestas ampulares las unidades de cc oscuras se encuentra adyacentes al planum semilunatum, por lo que muchos autores han confundido las cc oscuras con las del plano semilunatum. Son de menor altura que las transicionales y contiene numerosos microvelli en su superficie apical.
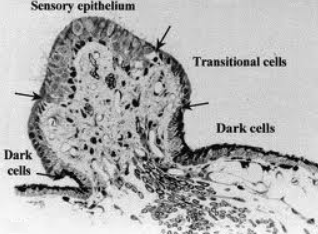 Las cc oscuras son esencialmente idénticas morfológica y funcionalmente a las cc marginales o superficiales de la estría vascular. Tienen forma cuboidea o aplanada y un citoplasma de carácter electrodenso. Su citoplasma contiene aparato de Golgi, mitocondrias, vesículas de micropinocitosis, cuerpos multivesiculares y un gran número de vacuolas, algunas de carácter autofágico. Posee fogosomas, o lisosomas secundarios, en los que pueden detectarse melanosomas pobremente pigmentados. En su superficie distal presenta prolongaciones e interdigitaciones que alojan mitocondrias y entre las cuales se ha podido detectar una actividad ATP-asa positiva lo que indica una participación activa de estas formaciones en los fenómenos de transporte iónico y de agua. También se ha sugerido que pueden tener una función de reabsorción y degradación otoconial al haberse observado en las cc oscuras próximas al utrículo algunas otoconias adheridas a su superficie.
Las cc oscuras son esencialmente idénticas morfológica y funcionalmente a las cc marginales o superficiales de la estría vascular. Tienen forma cuboidea o aplanada y un citoplasma de carácter electrodenso. Su citoplasma contiene aparato de Golgi, mitocondrias, vesículas de micropinocitosis, cuerpos multivesiculares y un gran número de vacuolas, algunas de carácter autofágico. Posee fogosomas, o lisosomas secundarios, en los que pueden detectarse melanosomas pobremente pigmentados. En su superficie distal presenta prolongaciones e interdigitaciones que alojan mitocondrias y entre las cuales se ha podido detectar una actividad ATP-asa positiva lo que indica una participación activa de estas formaciones en los fenómenos de transporte iónico y de agua. También se ha sugerido que pueden tener una función de reabsorción y degradación otoconial al haberse observado en las cc oscuras próximas al utrículo algunas otoconias adheridas a su superficie. Su núcleo está situado en el centro celular y puede ser lobulado.
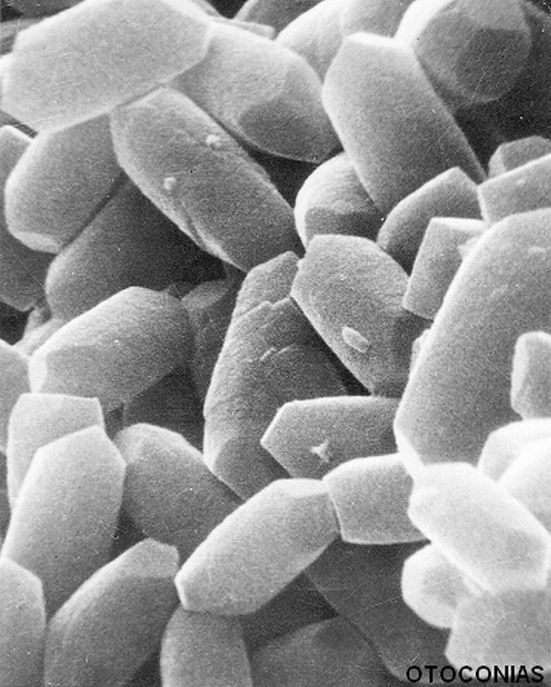 LOS OTOLITOS.
LOS OTOLITOS. Los otolitos son concreciones salinas de carbonato cálcico en forma de calcita, de tamaño variado, de unas diez micras y de elevado peso específico y, por ende, sensibles a la gravedad. Son elementos esenciales de los receptores maculares, utricular y sacular. Están anclados en la masa gelatinosa de la membrana otolítica mediante fibras de colágeno, pero pueden desprenderse y disolverse en el liquido del espacio endolinfático. El metabolismo del otolito adulto está sometido a un permanente intercambio de calcio a todo lo largo de su superficie externa.
Forma.
Se han descrito tres patrones morfológicos de otolitos maduros:
- El más habitual y abundante es el prismático hexagonal con dos extremos que poseen tres facetas romboédricas.
- Otra forma es la de una estructura multifacetada que recuerda la forma de diábolo.
- El tercer modelo se correspondería con la forma geométrica del romboedron (seis caras de rombos no cuadrados).
Tamaño.
Su longitud oscila entre 0,1 y 30 μm y tienen un elevado peso específico.
En la mácula utricular los cristales más grandes son los situados en situación lateral que es además donde forman una capa más espesa. Por el contrario, más medialmente, junto a la estriola, es donde los cristales son más pequeños y la capa de otolitos es más delgada.
Son duros y su estructura es muy homogénea situándose las zonas más densas en la periferia y las más claras en el centro y en las extremidades. Poseen un núcleo central a partir del cual se inicia la formación del cristal. Impiden el paso de electrones.
No está claro si están formados por un solo cristal o se trata de estructuras policristalinas.
Dinámica.
El metabolismo del otolito adulto está sometido a un permanente intercambio de calcio en toda su superficie externa.
Experimentan un ciclo dinámico de renovación permanente.
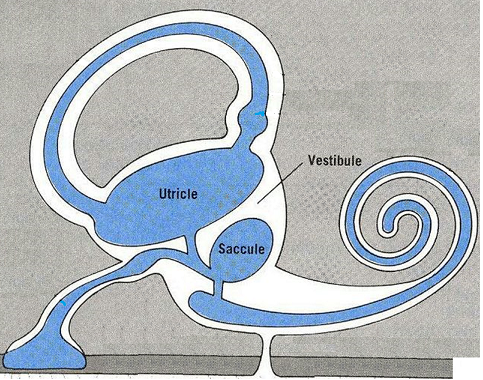 SACO ENDOLINFÁTICO.
SACO ENDOLINFÁTICO. El canal endolinfático se forma tras la unión en Y de dos conductos de corto recorrido nacidos uno en el utrículo y otro en el sáculo. El recorrido de este canal termina en un fondo ciego que es el saco endolinfático.
La histología del saco endolinfático es muy simple. Está constituido por una sola capa de cc epiteliales, epitelio pavimentoso o cúbico, que descansan sobre una membrana basal y un tejido conjuntivo laxo muy vascularizado rico en capilares fenestrado.
En la región proximal el epitelio es cuboideo. La altura y la anchura de las cc oscila aproximadamente entre 5 y 6 µm. Estas cc presentan microvellosidades en su superficie y vesículas de pinocitosis en su citoplasma como datos microscópicos más característicos.
La región intermedia o rugosa es denomina así por lo pliegues que presenta su superficie intraluminal. En esta zona las cc son prismáticas o columnares y presentan una altura de 10 a 20 µm y una anchura de 4 a 8 µm. Se distinguen dos tipos de cc, las claras y las oscuras.
Las claras presentan una superficie cupuliforme muy rica en microvellosidades. En el citoplasma poseen numerosas vesículas, vacuolas, mitocondrias y ribosomas libres. El aparato de Golgi y el retículo endoplásmico rugoso están poco desarrollados. El núcleo es ovoideo y se sitúa en el centro de la cc.
Las cc oscuras presentan en su superficie escasas microvellosidades, sus ribosomas, vesículas y mitocondrias son escasos y el citoplasma es muy electrodenso. El núcleo es central y suele presentar indentaciones. A este nivel las cc oscuras no tienen invaginaciones en sus membranas basolaterales. En la luz de esta región del saco existen sustancias electrodensas y macrófagos intraluminales con numerosos lisosomas en su citoplasma.
La porción distal está compuesta por cc columnares de una altura de 10 µm y una anchura de 5 µm. en esta región se han diferenciado también cc claras y cc oscuras. Las claras son más abundantes. Hacia el fondo del saco las cc disminuyen de altura y su superficie se hace más lisa.
Todas las cc oscuras del SE de las diferentes regiones están implicadas en el transporte iónico.
Las cc claras tienen una función secretora paracrina de hormona natriurética, también conocida como saccina, que incrementa la producción de endolinfa. A nivel del SE se han identificado receptores de péptidos natriuréticos.
Funcionalmente el SE participa en tres funciones básicas:
- Secreción y reabsorción de la endolinfa vinculada a la presencia en las cc epiteliales de transportadores iónicos y acuaporinas.
- Respuesta inmunitaria del oído interno.
- Eliminación de restos presentes en la endolinfa, producida por los macrófagos intraluminales y su función de fagocitosis.
Para continuar visualizando este capítulo descargar tema completo
