-
TÁLAMO.
-
SISTEMA NERVIOSO CENTRAL.
Las principales funciones que cumple el cerebelo son las coordinación y organización del movimiento. A través de sus conexiones recibe información motora elaborada en el nivel cortical y compara y corrige el desarrollo y evolución de los actos motores. También actúa reforzando el movimiento para mantener el tono muscular de acuerdo a los cambios de posición. Algunos pacientes con trastornos cerebelosos presentan patologías como la ataxia cerebelosa que se caracteriza por falta de coordinación del equilibrio, la marcha, el movimiento de las extremidades y de los ojos. Además estudios dicen que el cerebelo está involucrado al condicionamiento clásico de parpadeo y en el aprendizaje de secuencias motoras
El cerebelo es una de las partes fundamentales del sistema del equilibrio pues interviene en tres funciones principales:
- El cerebelo hace que los movimientos corporales sean sinergéticos, eumétricos y euérgicos. Estas son tres cualidades que han de tener los movimientos corporales para ser efectivos en el mantenimiento del equilibrio y en todas las reacciones motoras del cuerpo en general.
- Hace que en todo momento el cuerpo adopte una postura adecuada para mantener el equilibrio según su posición en el espacio.
Esta función exige de un aprendizaje. El cerebelo mediante una integración de la información sensorial que recibe tiene elaborados modelos o programas internos de secuencia y tipos de movimientos a realizar para el mantenimiento del equilibrio tanto en movimiento como de forma estática adoptado la postura adecuada a las necesidades del individuo en cada momento para mantener su equilibrio corporal. Complementando esta función tiene además la capacidad de realizar una función anticipatoria en cuanto al ajuste postural, con el objetivo principal de estabilizar el cuerpo en movimiento y ejercer una función anticipatoria en cuanto al ajuste postural, con el objetivo principal de estabilizar el cuerpo en movimiento y en reposo.
Neurofisiológicamente para realizar incluso los movimientos más sencillos es evidente que se han de realizar actos y contracciones complejas. Se requiere: la contracción de un grupo o grupos de músculos determinados, la relajación simultánea del conjunto o conjuntos de músculos antagonistas, un nivel específico de contracción muscular durante un periodo de tiempo preciso, y la secuenciación adecuada de la contracción y relajación de los grupos musculares necesarios para el movimiento en cuestión. El número de fibras musculares activadas determina el alcance de la contracción muscular. A su vez, el número de fibras musculares que se contraen en un momento dado (es decir, la fuerza o intensidad de la contracción) está en función del número de motoneuronas alfa que se activan. La duración de la contracción viene determinada, en gran medida, por la duración de la activación de las fibras nerviosas que inervan los músculos necesarios para el acto específico.
Estas dos funciones qie hemos descrito para los movimientos corporales son igualmente aplicables a la motricidad ocular. El cerebelo afina cada uno de los subtipos de movimientos oculares para que funcionen conjuntamente a fin de centrar y mantener imágenes de objetos de interés sobre la fóvea. El flóculo/paraflóculo están realacionados con el control de los movimientos oculares de seguimiento o persecución lenta y con los del mantenimiento de la mirada. El vermis oculomotor dorsal y la porción posterior del núcleo fastigial (la región oculomotora fastigial) están relacioandos con las sacadas y el inicio del seguimiento ocular.
- La tercera función es su participaciónen el mantenimiento del tono muscular.
El control que ejerce sobre las funciones motoras es siempre homolateral, es decir, que cada hemisferio cerebeloso controla la mitad correspondiente del organismo. Cave preguntarse como el cerebelo ejerce esta función de control sin tener el control directo sobre la contracción muscular: el cerebelo vigila y establece ajustes correctores de las actividades motoras desencadenadas por otras partes del encéfalo.
- La tercera función es su participaciónen el mantenimiento del tono muscular.
El control que ejerce sobre las funciones motoras es siempre homolateral, es decir, que cada hemisferio cerebeloso controla la mitad correspondiente del organismo. Cave preguntarse como el cerebelo ejerce esta función de control sin tener el control directo sobre la contracción muscular: el cerebelo vigila y establece ajustes correctores de las actividades motoras desencadenadas por otras partes del encéfalo.
Tanto para la ejecución precisa y eficaz de movimientos intencionados, así como para la presencia de una postura adecuada asociada a la bipedestación y al movimiento, el cerebelo es capaz de integrar y organizar la secuencia de acontecimientos asociados a la respuesta. Funciona como un ordenador para integrar señales sensoriales y motoras por sus propiedades de integración y de análisis de las señales aferentes y poseer las conexiones recíprocas para formar una serie de vías de "retroalimentación" hacia sus fuentes aferentes.
La corteza cerebelosa recibe continuamente información actualizada procedente de todos los elementos sensoriales del sistema del equilibrio: regiones del SNC relacionadas con la regulación de las funciones motoras, así como señales sensoriales directas e indirectas relacionadas con el estado de músculos individuales, grupos de músculos y otras señales que incluyen impulsos táctiles, aferencias cutáneas y aferencias articulares. De esta forma está informado de la posición corporal, ritmo de movimiento, las fuerzas que actúan sobre el cuerpo, etc. Además envía respuestas de "retroalimentación" a estas las regiones de las que recibe información.
Todas la conexiones nerviosas del cerebelo sale o entra por los pedúnculos cerebelosos.
Información recibida por el cerebelo.
Al cerebelo llegan aferencias de todas las vías motoras y de todas las sensitivas, incluyendo la olfatoria.
Todas las aferencia informativas recibidas por el cerebelo confluyen en el núcleo floculonodular y en el vermis. Ascienden a través de fibras musgosas y trepadoras hacia la corteza cerebelosa de Purkinje para excitarla.
El cerebelo recibe 40 veces más fibras aferentes que las eferentes que emite.
Las fibras musgosas están formadas por fibras terminales de todos los tractos aferentes cerebelosos, pudiendo una sola fibra estimular a miles de cc de Purkinje a través de las cc granulosas. Las fibras trepadoras son las fibras terminales de los tractos olivocerebelosos a través de las capas de la corteza y una sola fibra hace contacto con una sola cc de Purkinje.
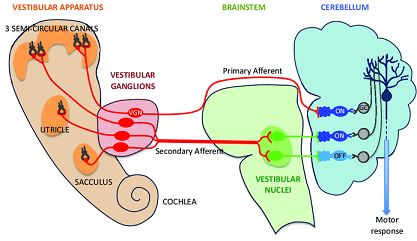 Información vestibular.
Información vestibular. Ver capítulo 02.3ª.06.
Información procedente de la médula espinal.
A nivel de la médula espinal las aferencias al cerebelo llegan a través de los tractos espinocerebelosos anterior y posterior que tienen su origen en las interneuronas de la sustancia gris espinal (columna de Clarke). Estos tractos son capaces de transmitir impulsos nerviosos más rápido que cualquier otra vía del SNC alcanzando una velocidad de 120 m/s (metros por segundo), siendo los tractos aferentes más rápidos de la médula. Esta rapidez es necesaria para que llegue al cerebelo la información sobre los cambios ocurridos en los grupos musculares periféricos y poder coordinarlos a tiempo. Recogen información de los husos neuromusculares, Órgano de Golgi, dolor e información cutánea para transmitirla hacia los núcleos cerebelosos. El vermis, a través de esto dos fascículos, recibe información de prácticamente todos los elementos motores que intervienen en el equilibrio.
Tracto espinocerebeloso anterior (ventral) o de Gowers se origina en la médula, en neuronas que asientan en la zona lateral de la base del asta posterior, entre los últimos segmentos lumbares y los sacrococcígeos. Algunas de sus fibras cruzan la comisura gris para ascender por el cordón lateral del lado contrario, donde se sitúa próximo a la superficie medular. Las pocas fibras que no se cruzan ascienden por el cordón lateral del mismo lado. Todas sus fibras atraviesan el bulbo y el puente, y llegan hasta la zona más caudal del mesencéfalo donde cambian bruscamente de dirección para entrar al cerebelo por el pedúnculo superior. Alcanza el vermis y las bandas paravermianas de ambos lados. Transporta información propioceptiva generada internamente sobre el ritmo locomotor, información propioceptiva de articulaciones, tendones (aferentes Ib) y ligamentos ante movimientos voluntarios, para poder realizar una comparación con los programas motores del cerebelo. Transmite además información propioceptiva inconsciente y exteroceptiva de la extremidad inferior.
Tracto espinocerebeloso posterior (dorsal) o de Flechsing está formado por axones de neuronas cuyo soma se localiza en la columna torácica o núcleo de Stilling-Clarke. Asciende por el cordón lateral pegado a la superficie y justo por detrás del tracto espinocerebeloso anterior. Al alcanzar el bulbo penetra en el cerebelo por el pedúnculo inferior y llega hasta el vermis y la banda paravermiana del mismo lado de su origen. Proporciona al cerebelo información detallada sobre la evolución de los movimientos (propiocepción inconsciente). La columna de Clarke recibe información propioceptiva del huso (aferente Ia), tanto de movimientos pasivos como voluntarios de manera ipsilateral. Transmite información propioceptiva inconsciente y exteroceptiva procedente del tronco y la extremidad inferior.
Información procedente del bulbo raquídeo.
Las aferencias a este nivel llegan por medio de los tractos cuneocerebeloso, olivocerebeloso y reticulocerebeloso.
Tracto cuneocerebeloso, está formado por los axones de las neuronas que asientan en el núcleo cuneiforme accesorio (fibras arqueadas externas posteriores). Asciende por el bulbo raquídeo e ingresa en el hemisferio cerebeloso del mismo lado a través del pedúnculo cerebeloso inferior mezclado con el tracto espinocerebeloso posterior. Acaba en el vermis y en la banda paravermiana del mismo lado. Emite algunas ramas colaterales que terminan en los núcleos profundos cerebelosos. Este tracto es portador de información muscular de los husos musculares y tendinosos y de los receptores articulares de la extremidad superior y la parte superior del tórax. Transmite la sensibilidad propioceptiva inconsciente y exteroceptiva de la mitad superior del cuerpo.
Tracto olivocerebeloso es la conexión más importante que se establece entre bulbo raquídeo y cerebelo. Está formado por axones de las neuronas del núcleo olivar inferior y de los núcleos olivares accesorios. Estos núcleos reciben información somatoestésica, visual y de la corteza cerebral además de recibir aferencias vestibulares y del propio cerebelo. Al poco de originarse, el tracto olivocerebeloso se decusa totalmente y entra en el cerebelo por el pedúnculo inferior. Termina proporcionando fibras trepadoras para toda la corteza cerebelosa. Transmite al cerebelo la información recibida por los núcleos olivares.
Tracto reticulocerebeloso está formado por axones de neuronas localizadas en la formación reticular bulbar y póntica. Parte de las fibras se cruzan y otra parte van directas. Entra por el pedúnculo cerebeloso inferior y alcanza principalmente el espinocerebelo aunque también manda algunas fibras para el cerebrocerebelo. Transmite información compleja, tanto de la periferia como de la corteza cerebral y otras partes del SNC.
Tracto Caeruleocerebeloso. El locus coeruleus es unos de los nucleos más pequeños pero con mas proyecciones a otras partes del sistema nervioso. Así emite un tracto que penetra por el pedúnculo cerebeloso superior y termina distribuido por todos los núcleos y la corteza del cerebelo.
Tracto pontocerebeloso. Además de los núcleos de los pares craneales, en el puente hay una gran cantidad de neuronas pontinas organizadas en numerosos núcleos pequeños llamados núcleos del puente. Este trato está formado por fibras que se originan de los núcleos del puente y siguen un trayecto horizontal por la protuberancia, se decusan y entran por el pedúnculo cerebeloso medio. Terminan en la corteza de los hemisferios y en el núcleo globoso.
Tracto Rafecerebeloso: se originan del núcleo del rafe a nivel de bulbo raquídeo e ingresan al cerebelo a través de los pedúnculos cerebelosos medios.
Información procedente del tálamo.
A nivel del mesencéfalo las aferencias llegan por medio de los tractos tectocerebeloso, trigeminocerebeloso y rubrocerebeloso.
Tracto tectocerebeloso está formado por los axones de las neuronas de los tubérculo cuadrigéminos superiores e inferiores. Entran en el cerebelo a través del pedúnculo superior del mismo lado y terminan en la parte media del vermis. Transmite información visual y acústica proveniente de la corteza cerebral.
Tracto trigeminocerebeloso está formado por axones de neuronas del núcleo mesencefálico del nervio trigémino que entran al cerebelo a través del pedúnculo superior sin decusarse por el camino. Terminan en el vermis y en la banda vermiana del mismo lado de su origen. Transmite información propioceptiva del macizo craneofacial.
Tracto rubrocerebeloso está formado por axones de neuronas asentadas la porción parvocelular del núcleo rojo que se decusan en su totalidad antes de alcanzar el cerebelo por el pedúnculo superior.
Información procedente del cerebro.
Todas las aferencias que recibe el cerebrocerebelo forman parte del tracto corticopontocerebeloso. Este tracto se origina en una amplia zona de la corteza cerebral que abarca los lóbulos frontal, parietal, occipital y temporal, y antes de entrar en el cerebelo hace sinapsis en los núcleos del puente. La mayoría de las fibras que van desde la corteza hacia los núcleos del puente son colaterales de axones que se dirigen hacia otras zonas del encéfalo o hacia la médula espinal y cuyo cuerpo neuronal se sitúa en la capa V del cortex cerebral. Estas fibras se pueden dividir, según su origen, en: frontopónticas, parietopónticas, occipitopónticas y temporopónticas.
Fibras frontopónticas se originan en las cortezas motora y premotora, y pasan por el brazo anterior de la cápsula interna. En el mesencéfalo, discurren por la base de los pedúnculos cerebrales medialmente al tracto corticonuclear. Terminan en los núcleos del puente más mediales.
Fibras parietopónticas se originan en las áreas somatosensitivas primaria y secundaria y en áreas visuales. Pasan por el brazo posterior de la cápsula interna y luego por la base de los pedúnculos cereberales lateralmente al tracto corticoespinal. Terminan en los núcleos del puente más laterales.
Fibras occipitopónticas se originan en áreas secundarias relacionadas con el procesamiento de estímulos visuales del movimiento (corriente magnocelular de la vía óptica). Pasan por la porción retrolenticular de la cápsula interna y luego por la base de los pedúnculos cereberales lateralmente al tracto corticoespinal. Terminan en los núcleos del puente más laterales. Estas fibras proporcionan informacion del sistema visual al cerebelo.
Fibras temporopónticas pasan por la porción sublenticular de la cápsula interna y a nivel del mesencéfalo se colocan lateralmente al tracto corticoespinal. Termina en los núcleos del puente más laterales.
Las fibras que van desde los núcleos del puente al cerebelo (fibras pontocerebelosas) siguen un trayecto horizontal por la protuberancia, se decusan y entran por el pedúnculo medio. Terminan en la corteza de los hemisferios y en el núcleo globoso.
Toda esta información recibida se va procesando y permite al floculonodular y a la úvula del vermis elaborar una representación tridimensional de los movimientos corporales que será utilizada para ser comparada con la que tiene almacenada. Es decir que compara la información que recibe en cada momento sobre el estado del equilibrio con la que tiene almacenada. Si las dos informaciones, la almacenada y la recibida, no son concordantes, de manera instantánea se trasmiten señales correctoras adecuadas hacia el sistema motor para aumentar o disminuir la actividad de músculos específicos. Esta función rectora se ejerce tanto sobre la actividad refleja como sobre la voluntaria.
Información enviada desde el cerebelo.
Del cerebelo parten eferencias para controlar todas las vías motoras descendentes.
Todas las eferencias de la corteza cerebelosa se realizan a través de las cc de Purkinje y terminan haciendo sinapsis con las neuronas de los núcleos profundos del cerebelo que constituyen la vía eferente fundamental del cerebelo. Las eferencias no suelen hacer sinapsis directamente sobre las motoneuronas de la vía final común excepto en las de los músculos extrínsecos del globo ocular. Las eferencias normalmente actúan sobre los núcleos motores del tronco del encéfalo. Estas fibras eferentes cerebelosas se conectan con el núcleo rojo, el tálamo, el complejo vestibular y la formación reticular.
Información remitida a la corteza cerebral.
Se realiza por la vía dentotalámica. Los axones de las neuronas del núcleo dentado (núcleo profundo cerebeloso) discurren a través del pedúnculo cerebeloso superior y cruzan la línea media hasta el otro lado en la decusación del pedúnculo cerebeloso superior. Las fibras terminan haciendo sinapsis con células en el núcleo ventrolateral del tálamo contralateral. Los axones de las neuronas talámicas ascienden a través de la cápsula interna y la corona radiada y terminan en el área motora primaria de la corteza cerebral contralateral. A través de esta vía el cerebelo envía información aferente hacia las áreas motoras del córtex cerebral para ajustar la postura y llevar a cabo la “corrección de errores”.
El tálamo integra la información sensorial antes de llegar a la corteza cerebral. Por este motivo existen pacientes atáxicos con lesiones talámicas, su cerebelo está intacto, pero su tálamo no integra bien la información.
Información remitida a núcleos basales.
Los ganglios basales son agrupaciones de neuronas subcorticales que se encuentran localizadas en las profundidades del encéfalo. Se incluyen los siguientes: Núcleo caudado (una estructura en forma de C que se estrecha en una cola delgada), Putamen, Globo pálido (situado al lado del putamen), Núcleo subtalámico, Sustancia negra. En estos ganglios basales mediante las conexiones que existen entre sus distintas estructuras se decide qué movimientos son los que se realizan y cuales no, ayudan a iniciar y suavizar los movimientos musculares, suprimir los movimientos involuntarios y coordinar los cambios de postura.
El cerebelo tiene conexiones directas a los ganglios basales a través del núcleo dentado. Esta información participa en el ajuste fino necesario para la destreza manual. Debemos recordar que tanto el circuito de los ganglios basales como el circuito cerebeloso son bucles subcorticales que en gran medida reciben información de la corteza y la devuelven a través del tálamo. La lesión en esta vía puede dar signos como la distonía.
Información remitida al troncoencéfalo.
Tracto interpuestorreticular se origina en el núcleo interpuesto del cerebelo (formado por el núcleo globoso y el emboliforme), sus fibras se decusan parcialmente y salen del cerebelo por los pedúnculos inferiores para alcanzar los núcleos de la formación reticular.
Tracto interpuestoolivar sale por el pedúnculo cerebeloso superior, se decusa en su totalidad a nivel del mesencéfalo y desciende por el tronco del encéfalo para alcanzar el núcleo olivar inferior.
Tracto interpuestotectal se decusa parcialmente antes de salir por el pedúnculo cerebeloso superior y ascender por el tronco del encéfalo hasta alcanzar los tubérculos cuadrigéminos superior e inferiores.
Tracto interpuestorrúbrico (Cerebelorubral), también denominado vía globoso-emboliforme-rúbrica es la eferencia cerebelosa más importante del y principal vía de descarga del núcleo interpuesto. Los axones de las neuronas de los núcleos globoso y emboliforme (núcleos profundos del cerebelo que forman el interpuesto) discurren a través del pedúnculo cerebeloso superior y cruzan la línea media en el mesencéfalo y alcanzan el núcleo rojo contralateral. Desde el núcleo rojo parten axones hacia el núcleo ventral intermedio del tálamo que, a su vez, envía axones para la corteza cerebral motora y sensorial. Controla la actividad de las vías motoras homolaterales que descienden hasta la médula espinal.
Via fastigiorreticular. Los axones de las neuronas del núcleo fastigio discurren a través del pedúnculo cerebeloso inferior y terminan haciendo sinapsis con neuronas de la formación reticular.
Tracto floculoculomotor: se origina en los flóculos, se decusa en pleno cerebelo, sale por el pedúnculo superior y asciende por el tronco del encéfalo hasta llegar al núcleo del nervio oculomotor (o motor ocular común). Controla los movimientos del globo ocular
Información remitida a los núcleos vestibulares.
Vía fastigiovestibular (cerebelovestibular). Los axones de las neuronas del núcleo fastigio discurren por el tracto cerebelovestibular a través del pedúnculo cerebeloso inferior y terminan proyectándose sobre las neuronas del núcleo vestibular lateral y medial de ambos lados. Debe recordarse que algunos axones de las cc de Purkinje se proyectan directamente hasta el núcleo vestibular lateral. Las neuronas del núcleo vestibular lateral forman el tracto vestibuloespinal y esta vía nerviosa fastigio vestibular tiene como función regular la actividad de los tractos vestibuloespinales medial y lateral.
Neurológicamente las aferencias y eferencias cerebelosas constituyen un circuito retículo-cerebelo-vestibular que forma una unidad funcional al servicio del sistema del equilibrio y la locomoción, actuando sobre la vía final común de la médula espinal para ejecutar respuestas reflejas posturales en el sentido de modular, afinar, matizar y coordinar las coordenadas de los comandos motores, es decir, hace que los movimientos osteomusculares tengan precisión. Además, al ser el órgano de gobierno de los circuitos propioceptivos supramedulares inconscientes reflejos, ejerce a través de ellos una regulación del tono muscular en los músculos agonistas y antagonistas.
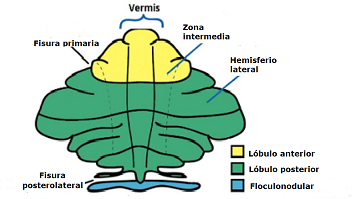
Topografía funcional del cerebelo.
Nódulo floculonodular.
También conocido como vestibulocerebelo, es el área funcional mas importante en el control del equilibrio y en la regulación del reflejo óculo-vestibular. Transporta información sobre la posición de la cabeza y el cuerpo en el espacio, y señales útiles para orientar los ojos durante los movimientos.
Recibe información de los órganos vestibulares ipsilaterales para enviarla a la formación reticular.
Ejerce su acción sobre la postura y el equilibrio mediante proyecciones vestibuloespinales y reticuloespinales a las motoneuronas extensoras que actúan sobre los músculos axiales y proximales de las extremidades.
Vérmis.
También conocido como espinocerebelo, su corteza se encarga del control de la postura y los movimientos del eje mayor del cuerpo, en particular el equilibrio del tronco y la marcha (equilibrio axial) realizando los reajustes posturales y las reacciones anticipatorias.
Las zonas laterales al vermis, que se consideran como zona intermedia del hemisferio cerebeloso, reciben información sobre la posición y movimiento de las extremidades a través de la información propioceptiva y cutánea que recibe de la médula espinal y controla los músculos de las partes dístales de las extremidades.
Recibe información sensitiva ascendente (tracto espinocerebeloso), la cual llega a la corteza del cerebelo a través de las fibras musgosas y trepadoras. Esta información es comparada y corregida para que, a través de vías descendentes, se transmita un movimiento cada vez más fino y preciso, ajustándose cada vez más a las demandas ambientales.
Hemisferios cerebelosos.
Esta área funcional del cerebelo lleva a cabo tanto la planificación del movimiento voluntario como la temporalización o “timing”, o secuencialización del movimiento y el inicio motor en todo el cuerpo. Especialmente del brazo, el antebrazo y la mano y sincronización de dichos movimientos.
Las aferencias llegan del cerebro, más concretamente de la corteza. El movimiento planeado viaja hacia el cerebelo, y más tarde a el tronco del encéfalo y el tálamo, volviendo de nuevo a la corteza motora y premotora. De esta manera se modulan la actividad de las neuronas corticales que se proyectan a la médula espinal contralateral.
Con respecto al SOM son funciones específicas del cerebelo:
- El mantenimiento de posiciones excéntricas de la mirada.
- La producción de los movimientos de seguimiento.
- Modulación de la amplitud de las sacadas, haciendo que estas se ajusten funcionalmente a los movimientos cefálicos.
Como dijimos en la anatomía del sistema del equilibrio, a nivel troncoencefálico se encuentran toda una serie de núcleos que son fundamentales para el Sistema del Equilibrio y que junto con el cerebelo conforman un sistema de control y regulación del mismo. Están ubicados a lo largo del FLM: núcleos vestibulares, oculomotores etc. Aquí nos referiremos a la sustancia reticular, al núcleo de la oliva inferior y a los ganglios basales.
 Sustancia Reticular.
Sustancia Reticular. También llamada sistema centroencefálico de Panfield o formación reticular.
Está constituida por una compleja red de núcleos (más de 100) y neuronas situados a lo lago del tronco encefálico que sirve como centro principal de integración y transmisión de muchos sistemas cerebrales vitales. algunas de sus funciones de coordinación son necesarias para la supervivencia.
En las preparaciones histológicas las fibras junto con los núcleos la dan un aspecto de red y de ahí el nombre de reticular.
Anatómicamente no presenta límites definidos, y los numerosos núcleos incluidos en esta estructura no tienen delimitaciones territoriales precisas. Se extiende a lo largo del bulbo, protuberancia y mesencéfalo, alcanzando finalmente algunos sectores del tálamo óptico.
Funcionalmente constituye una formación de interneuronas multisinápticas entre las vías que conectan la medula espinal y el cerebro siendo el mayor centro integrador de todo el SNC. Además de constituir un gran centro de transmisión entre varios sistemas del SNC, tiene una función de integrar, coordinar e influir en varias regiones tanto del SNC como del periférico, realizando funciones necesarias para la supervivencia. Esta región es capaz de modificar la actividad neuronal de las fuentes de estímulos que recibe y de suprimir o amplificar la excitabilidad de muchas neuronas. También, aunque en menor grado, está capacitada para emitir impulsos generados en ella misma sin ser respuestas a una previa información periférica.
Las funciones más importantes de la sustancia reticular son:
- Función de vigilia sueño.
- Control del dolo.
- Regulación de la actividad visceral.
- Control de los movimientos voluntarios.
Con respecto al equilibrio la función que nos interesa analizar es el control de los movimientos voluntarios. Esta función es vehiculada por los siguientes tractos cómo los más importantes:
- El Tracto Retículo-Espinal, que se integra en los otros tractos descendentes para la postura.
- El Tracto Vestíbulo-Espinal.
- El Tracto Córtico-Espinal o vía piramidal.
Tanto el retículo-espinal como el vestíbulo-espinal actúan sobre la postura y sobre la musculatura axial y proximal de las extremidades, fijando y moviendo las grandes articulaciones. La vía piramidal es la que controla los movimientos finos.
Para el control de los movimientos el estímulo parte de los núcleos situados en la Protuberancia o Puente de Varolio que son productores de glutamato y pertenecen al grupo de los núcleos centrales. Algún núcleo situado en Bulbo Raquídeo también participa en esta función, así como alguno del grupo central.
Las fibras de proyección descienden tanto de forma directa como cruzada, ya sea por el cordón anterior o por el lateral de la médula espinal. Los que provienen de la protuberancia descienden por el cordón anterior y las de los núcleos del grupo central lo hacen más por los cordones laterales. Actúan en las astas anteriores de la medula sobre las motoneuronas alfa de los de la musculatura axial y proximal de las extremidades.
Las fibras procedentes de la protuberancia actúan sobre todo en la musculatura extensora. Cuando el paciente sufre una descerebración donde la lesión esta a nivel del mesencéfalo, en el núcleo Rojo o por debajo, se libera la acción de estos núcleos, activando la musculatura extensora. Esta será la respuesta a la descerebración. Estos núcleos reciben fibras: de la corteza cerebral (fibras Cortico-Reticulares), de el núcleo pedúnculo pontino colinérgico, del cerebelo y de la médula espinal por la vía Espino-Reticular.
Recibe información de todas las vías sensitivas y sensoriales, encontrándose en una estrecha relación con todos los circuitos del sistema nervioso y en particular del equilibrio, tanto ascendentes como descendentes. Su función es es hacer de seleccionador tanto de los impulsos informativos que recibe como de los eferente que esta formación emite, dando prioridad de paso a aquellos que realmente deben de predominar en cada momento por su significado. Es decir, que recibe impulsos, que tras integrarlos, puede transmitirlos, o no. De este modo filtra la información y emite impulsos que pueden ser ascendentes hacia el cerebelo y corteza cerebral, o descendentes a los núcleos estriomotores de la sustancia gris de la médula espinal y muy especialmente a las motoneuronas gamma.
Con respecto a su función en el sistema del equilibrio, lo que tiene mayor interés, son sus mensajes descendentes. Mediante ellos ejerce el control permanente sobre los reflejos espinales segmentarios del tono muscular a través del circuito gamma (principalmente sobre los músculos extensores), adaptándolos a las necesidades posturales de cada momento.
La porción más caudal de la formación reticular, situada en el bulbo, recibe impulsos inhibitorios por parte de la corteza cerebelosa y cerebral que discurrirán por la vía córtico-retículo-espinal hasta la médula. De esta forma los mecanismos centrales relacionados con el control postural ejercen su influencia pasando antes por la Formación Reticular.
La porción cráneo-dorsal de la Formación Reticular tiene el significado contrario, es decir, es facilitadora de impulsos y recibe fibras del SV a través del núcleo de Deiters.
Merced a estos impulsos facilitadores e inhibitorios mantiene y prepara a los músculos para cualquier variación del tono exigida en cada momento por las circunstancias extrínsecas o intrínsecas.
Otra función de menor importancia es el control de las respuestas vegetativas al estímulo vestibular, ya que las conexiones entre el SV y los núcleos del sistema autónomo se realizan a través de la Formación Reticular.
Núcleo de la oliva inferior.
El cuerpo olivar u oliva bulbar consta de dos partes: el núcleo olivar superior y el núcleo olivar inferior.
El núcleo olivar inferior se divide a su vez en 3 núcleos principales por lo que también se conoce como complejo olivar inferior: núcleo olivar primario, núcleo olivar accesorio medial y el núcleo olivar accesorio dorsal.
Constituye un centro de convergencia e integración de información somatosensitiva, visual y de la corteza cerebral.
Sus eferencias se dirigen como fibras trepadoras que trasportarán información al cerebelo, donde harán sinapsis con las células de Purkinje, formando parte de un circuito de retroalimentación en el control y regulación de la actividad cerebelosa.
Ganglios basales.
En realidad no son ganglios sino núcleos grises subcorticales del diencéfalo. Intervienen en funciones esqueletomotoras, oculomotoras e incluso cognitivas y emocionales.
Funciones de los ganglios basales:
- Planificación, integración y control del movimiento voluntario.
- Actuación sobre el aprendizaje de las estrategias postrales y en la automatización de la conducta equilibratoria.
- Participación activa en las funciones mecánicas de la equilibracion corporal.
- Participación en conducta emocional y motivacional; algunos ganglios basales como el núcleo accumbens tienen conexiones con el sistema límbico y el sistema de recompensa cerebral, habida cuenta de su importancia en la gestión de la dopamina.
Participan en los circuitos subcorticales relacionados con el control de la postura y del movimiento, los sistemas esqueletomotor y oculomotor. Estos circuitos están muy separados unos de otros tanto estructural como funcionalmente, de tal forma que cada uno de ellos se origina en un área específica de la corteza cerebral e implica a diferentes partes de los ganglios basales y del tálamo.
Ahora bien no existen vías descendentes directas entre los ganglios basales y la medula espinal. Esto implica que el control motor de los ganglios basales es mediado a través de la Corteza Cerebral. Este control se lleva a cabo mediante vías ascendentes que desde los Ganglios Basales pasan por el Tálamo y llegan a la Corteza. El ganglio basal no inicia el movimiento sino la corteza cerebral.
El circuito esqueletomotor comienza y termina en las zonas motoras precentrales (la corteza premotora, el área motora suplementaria y la corteza motora) e implica tanto a las áreas somatosensitivas poscentrales de la corteza cerebral como al tálamo y a los ganglios basales, concretamente al núcleo putamen (núcleo de entrada), al globo pálido y la sustancia negra (núcleos de salida). Señalar que la información trasmitida por la corteza mantiene su organización somatotópica en el tálamo y en los núcleos basales. Otra cosa que parece clara es que los ganglios basales intervienen tanto regulando el movimiento propiamente dicho, es decir, una vez que éste se ha iniciado, como en la preparación del mismo (feedforward) y en el bucle de retroacción (feedback).
La conexión entre los ganglios basales y el tálamo se hace como dijimos a través de vías directas e indirectas. Las vías directas funcionan por retroalimentación positiva, es decir, la ausencia de señal es lo que permite que se activen las neuronas talámicas aumentando su actividad talamo-cortical y por tanto facilitando el movimiento. Por el contrario, las vías indirectas funcionan por una retroalimentación negativa, esto es, las eferencias de los ganglios basales inhiben a las neuronas del tálamo y por lo tanto el movimiento. Un aumento de la actividad en la vía indirecta tendría como resultados trastornos hipocinéticos (acinesia, bradicinesia, hipertonía, temblor de reposo... como sucede por ejemplo en la enfermedad de Parkinson), mientras que una disminución en la actividad de dicha vía ocasionaría trastornos hipercinéticos (discinesia, hipotonía, atetosis, posturas anormales mantenidas... como sucede en la corea o el balismo).
Otro circuito en el que participan los ganglios basales es en el circuito oculomotor, originado en los campos motores frontales y suplementario y que se proyecta al núcleo caudado desde el que parten proyecciones a la sustancia negra regresando a los campos oculares frontales y al tubérculo cuadrigémino superior con señales inhibitorias.
TÁLAMO.
Pero el tálamo fisiológicamente no es solamente un relais de paso hacia el córtex sino que además es un centro de procesamiento en el control y en la modulación de la trasmisión sensitiva de tal forma que, el tálamo determina si la información sensitiva alcanza o no la consciencia en la neocorteza. En esta función intervienen múltiples núcleos talámicos, como el complejo ventroposterior, el complejo ventroanterior-ventrolateral, los núcleos intralaminares y el grupo nuclear posterior (núcleos geniculados medial y lateral, pulvinar). Estos núcleos contienen neuronas multisensoriales que reciben entradas convergentes vestibulares, visuales y somatosensoriales que procesan y luego transmiten señales vestibulares, propioceptivas y visuales a la corteza vestibular.
Además el tálamo (concretamente los núcleos ventrolaterales del mismo) participa en la integración de la información motora del cerebelo y los ganglios basales y trasmite dicha información a las regiones de los hemisferios cerebrales que se ocupan del movimiento.
Al igual que otras zonas del cerebro el tálamo presenta su propio sistema de regulación o retroacción inhibitorias para regular su actividad y función.
SISTEMA NERVIOSO CENTRAL.
El SNC es el destino final de las múltiples aferencias sensoriales procedentes de todos los elementos que constituyen el sistema del equilibrio. Constituye un centro de procesamiento rector-codificador, que a partir de la información continuamente recibida, genera respuestas motoras siguiendo una serie de estrategias posturales que tiene configuradas para la regulación y control de la postura, del equilibrio y del movimiento corporal.
El córtex cerebral, además de controlar el equilibrio y el movimiento corporal, mediante la integración de la información recibida proporciona una percepción orientada de nuestro entorno inmediato, haciendo que el individuo esté orientado temporoespacialmente. No obstante, conseguir una buena orientación espacial es difícil, incluso en situaciones simples y comunes, ya que el sistema de interpretación e integración de la información es tan complejo que puede elaborar una síntesis inexacta en un momento dado, incluso en un individuo sano. Para comprender esto siempre se recurre al ejemplo del tren parado y la sensación de movimiento que se siente cuando otro tren paralelo se mueve.
Las regiones corticales que intervienen en la función equilibratoria del SNC son principalmente las áreas premotoras, motoras y somatoestésicas, como ya se explicó en la anatomía del equilibrio, capítulo 02.3ª.06. Estas áreas cerebrales, a su vez, se interconectan entre sí por intrincadas redes neuronales.
Anatómicamente, hasta el momento actual, se han identificado cuatro áreas: el área parietal 2V, situada en profundidad en la punta del surco intraparietal, el área 3aV localizada en el fondo del área somatoestésica primaria, una tercera situada en la parte posterior de la insula o córtex parieto-insular, y la cuarta denominada área MST que se corresponde con el área visual temporal medial y superior y con una región del lóbulo parietal inferior que es el área 7ª.
Para relizar el SNC su funcion procesadora y rectora-codificadora del Sistema del Equilibrio se realiza medida por una organización central del control del equilibrio basándose en cuatro elementos:
- Un valor de referencia estabilizado: es el punto de proyección en el suelo del centro de gravedad en condiciones estáticas.
- Señales detectoras de error: aquella información aferente proveniente del sistema laberíntico, visual, propioceptivo y cutáneo respecto a las situaciones de desequilibrios.
- Esquema corporal postural: aquel que informa sobre la orientación del cuerpo con respecto a la vertical gravitaria (receptores vestibulares, graviceptores somáticos), sobre la posición de los segmentos corporales unos respecto a otros (aferencias Ia de los husos musculares) y sobre sus propiedades dinámicas (sobre todo de las condiciones de apoyo).
- Reacciones posturales: aquellas que mantienen la posición de referencia y que se organizan a partir de los mensajes de error a través de dos tipos de bucles: uno continuo ante los cambios lentos de posición, y otro discontinuo y fásico que asegura una rápida corrección.
Las neuronas de las áreas corticales que intervienen en la función equilibratoria responden a los diversos tipos de estímulos sensoriales recibidos mediante la generación de un conjunto de respuestas posturales que se conocen con el nombre de estrategias de estabilización o ajustes posturales. El SNC participa ejerciendo un control en todas las estrategias posturales, sean reflejas o voluntarias.
Estrategias posturales.
Para compensar el efecto perturbador del movimiento y con la finalidad de prevenir o reducir al máximo el desplazamiento del centro de gravedad, el cerebro utiliza dos procedimientos. Uno de ellos implicaría un ajuste postural reaccional (reflejos), con el inconveniente de sobrevenir un cierto retraso ya que requieren información sensorial (feedback), mientras que el otro consistiría en una serie de ajustes posturales anticipados, que a través de redes nerviosas adaptativas, se liberarían como un todo antes de iniciarse la perturbación, están generadas centralmente (feedforward) y no requieren aferencias sensoriales.
Estas reacciones posturales pueden materializarse en la combinación de funciones motoras de supresión, habituación o anticipación para contrarrestar efectos indeseados, de calibración de impulsos sensitivos y estabilización inespecífica. Por ejemplo, el componente lento del nistagmo es admitido que tiene un origen laberíntico, pero su coordinación está controlada por estructuras centrales supranucleares.
Estas estrategias de estabilización son de tres tipos:
- Ajustes posturales reaccionales: reflejos.
Comprenden un conjunto de reflejos que, requiriendo información sensorial, contribuyen a mantener la postura: reflejos vestibuloespinales, reflejos vestibulocervicales, reflejos cervicocervicales, reflejos cervicoespinales, reflejos vestibulooculares, reflejos optocinéticos.
La finalidad de los reflejos posturales es mantener la postura de referencia o adaptarla al entorno, de tal manera que estos estabilizan en primer lugar la cabeza con respecto al espacio (vestibulocervicales y cervicocervicales), después el tronco en relación con la cabeza (vestibuloespinales, y cervicoespinales) y, finalmente, los miembros con relación al tronco.
El reflejo vestibulocervical es de gran importancia para estabilizar la cabeza con relación al espacio. Ante un movimiento cefálico la señal vestibular generada desencadena el reflejo vestibulocervical que actúa provocando una respuesta motora sobre los músculos del cuello para estabilizar la cabeza con relación al espacio oponiéndose al movimiento perturbador, lo que necesariamente tiende a anular la señal vestibular en su origen, es decir, este sistema funciona por retroacción negativa activado por el error.
El reflejo cervicocervical, por el contrario, responde a las señales propioceptivas del estiramiento de los músculos y de las articulaciones del cuello y genera una respuesta encargada de estabilizar la cabeza con relación al tronco (más que al espacio). En consecuencia, cuando el tronco está estacionario los reflejos vestibulocervicales y cervicocervicales colaboran en la estabilización de la cabeza. Pero cuando el tronco gira en relación con la cabeza, como cuando se mira a un objeto fijo al tiempo que se gira en una esquina el reflejo cervicocervical tiene que ser suprimido, de forma que el reflejo estibulocervical estabiliza por sí solo la cabeza.
La estabilidad postural se completa gracias a los reflejos vestibuloespinales y cervicoespinales, que funcionando de forma similar a los anteriores se complementan y se retroalimentan entre sí. Estos reflejos son los responsables, por ejemplo, de que en un cuadrúpedo cuando se le inclina la cabeza y el tronco hacia un lado, las respuestas vestibuloespinales son las responsables de extender las extremidades de dicho lado y flexionar las del lado contrario para oponerse a la perturbación. Ahora bien, si únicamente se inclina el tronco mientras que la cabeza permanece estacionaria respecto a éste, la respuesta cervicoespinal se opone a la inclinación. Sin embargo, si se hace girar la cabeza mientras el tronco permanece inmóvil, estos dos reflejos se oponen entre sí. Otra respuesta vestibuloespinal innata es la que se produce en las caídas repentinas, en las que el ser humano ofrece inesperadamente una respuesta extensora esteriotipada de los músculos antigravitatorios de las extremidades inferiores que sirven para evitar la caída, ahora bien, teniendo en cuenta que la latencia de la respuesta fija es inferior a 100 ms, la respuesta sólo es útil en caso de caídas desde alturas suficientemente grandes como para permitir que la respuesta tenga tiempo de presentarse. La contribución vestibular al control postural depende del tipo de perturbación impuesta y del lugar en el que se produce (pie, cabeza...). Todos estos reflejos, aún siendo innatos, pueden ser sometidos voluntariamente a un control superior para ajustarse al patrón intencional del movimiento voluntario con el fin de adaptarse al contexto funcional.
Los reflejos vestibulooculares son los responsables de mantener quietos los ojos cuando la cabeza se mueve con el fin de mantener sin movimiento las imágenes en la retina. Existen tres reflejos vestibulares diferentes, originados en los tres componentes principales del laberinto: 1. El reflejo vestibular de rotación, el cual compensa la rotación de la cabeza y recibe su aferencia predominantemente de los conductos semicirculares. 2. El reflejo vestibular de traslación, que compensa el movimiento lineal de la cabeza. 3. La respuesta ocular de contrarrotación, que compensa la inclinación de la cabeza en el plano vertical. Este reflejo, junto al anterior, reciben sus aferencias predominantemente desde los órganos otolíticos.
Cuando los conductos semicirculares perciben una rotación de cabeza en una dirección, los ojos rotan lentamente en la dirección opuesta para que la visión sea clara, si bien, posteriormente los ojos realizan un movimiento retrógrado a través del centro de la mirada originando un patrón repetitivo que recibe el nombre de nistagmo (combinación de fase lenta y rápido del movimiento ocular).
El movimiento lineal es percibido por los órganos otolíticos y es más complejo que el movimiento de rotación, ya que cuando la cabeza se mueve la imagen de un objeto próximo se desplaza más rápidamente en la retina que la de un objeto distante, de tal manera que el reflejo vestibular de traslación debe tener en cuenta la distancia a la que se encuentra el objeto visualizado, ya que cuanto mayor sea la distancia menor es el movimiento ocular. Los órganos otolíticos también perciben la aceleración lineal constante que la gravedad ejerce sobre la cabeza.
Ahora bien, los reflejos vestibulares precisan de la información del sistema optocinético para completar su función de estabilización de los ojos ya que en determinadas situaciones como es a la oscuridad o a los movimientos muy lentos, los órganos vestibulares no responden muy bien y precisan de los reflejos optocinéticos para completar su función.
Los reflejos optocinéticos proporcionan información al sistema vestibular, que éste utiliza para estabilizar los ojos. El hecho de que tanto el movimiento de imágenes en la retina, como el movimiento de cabeza induzcan al nistagmo y la percepción de movimiento, se debe a que las neuronas relacionadas con la visión (neuronas retinianas) se proyectan en los núcleos vestibulares en el mismo lugar al que llegan las señales vestibulares. Las neuronas que reciben aferencias no pueden distinguir entre señales visuales y vestibulares de manera que responden idénticamente al movimiento de una imagen en la retina, y probablemente esta es la razón por la que las personas no puedan distinguir en ocasiones entre ambos tipos de movimientos. Las células del núcleo de la cintilla óptica (neuronas retinianas) responden preferentemente a los estímulos que se mueven a través de la retina en una dirección temporal-nasal y a los estímulos cuya velocidad de movimiento es baja. Si bien, en los primates el reflejo optocinético está completado por un sistema cortical, que responde además a estímulos que se mueven a velocidades mayores o en una dirección nasaltemporal.
- Ajustes posturales anticipados.
Son movimiento para alcanzar posturas que prevén el efecto del movimiento con el fin de reducir al mínimo la alteración postural producida por este. Estos ajustes posturales aparecen ante la ejecución de los movimientos voluntarios, estabilizan la postura y se caracterizan por la contracción de ciertos grupos musculares que no intervienen directamente en el movimiento voluntario propiamente dicho. La orden central para un movimiento voluntario de una parte del cuerpo se asocia con una orden simultánea de acción anticipadora que prevé la perturbación postural esperada, y aunque los elementos básicos del control postural son innatos, es posible modificarlos de manera considerable mediante el aprendizaje. Si bien, los ajustes ante las alteraciones no esperadas dependen de la retroacción. Algunos de estos ajustes pueden ser relativamente rápidos y sencillos, como el reflejo miotático, pero por lo general son el producto de complejas reacciones motoras que se aprenden y se liberan como un todo. La magnitud y el tiempo de esos ajustes están relacionados con el contexto y con el movimiento realizado.
Estos ajustes posturales requieren la adquisición de información, por medio de la experiencia, y el aprendizaje y almacenaje de esas reacciones o sinergias que, de una forma prácticamente automática e inconsciente, se liberan como un todo.
Dentro de los ajustes posturales tiene especial relevancia en el mantenimiento de la postura bípeda el aprendizaje motor del control vestibuloocular, en el que interviene de una manera muy importante, al igual que en el resto de los ajustes posturales, el cerebelo. El aprendizaje motor del sistema vestibuloocular es adaptativo, ya que es capaz de ajustarse a la nueva demanda funcional, y plástico, ya que una vez establecido un cambio este se mantiene. Este cambio o ajuste en la eficacia sináptica recibe el nombre de ganancia nerviosa en la vía refleja.
Funciones equilibratorias del SNC
Las funciones equilibratorias del SNC se pueden resumir:
▪ Hace que el individuo tenga conciencia témporoespacial, es decir se sienta orientado, pudiendo sentirse equilibrado o desequilibrado. La orientación espacial es una capacidad fundamental para cualquier animal y es un requisito previo para el control postural, la locomoción y la relación con el ambiente. Permite tener la percepción, o sensación consciente, de los movimientos y actitud corporal. La conciencia temporoespacial se ubica en la circunvolución parietal ascendente siendo realizada por SNC mediante la capacidad que tiene de interpretación e integración de cada señal recibida para elaborar una síntesis informativa.
▪ El SNC mediante su capacidad de realiza una interpretación rápida y fiable de la información multisensorial que recibe va elaborando mediante el adecuado aprendizaje unos esquemas sensoriomotores a aplicar en cada situación según el equilibrio lo requiera. No obstante, en determinadas situaciones (como la cinetosis por ejemplo) no puede prevenir que se produzcan interpretaciones erróneas de la orientación espacial. Lo que esto significa es que la orientación espacial es una tarea difícil a pesar de la información multisensorial recibida y del aprendizaje.
▪ Controla todos los centros del equilibrio coordinándolos, modificando su actividad en sentido positivo o negativo. Mediante este control interviene en la regulación y control del tono corporal.
▪ Desarrollo de mecanismos de adaptación-compensación correctores de las desviaciones corporales producidas a partir de la última postura deseada, tanto en reposo, como durante el movimiento corporal. La interpretación rápida y fiable de toda la información multisensorial que recibe necesita un aprendizaje, pero todavía es mal conocido como el cerebro realiza la organización de mecanismos y esquemas neurológicos por los que regula el equilibrio y como los almacena. Es conocido que estos mecanismos de adaptación central son los que permiten al sistema del equilibrio autorregularse y autocorregirse, ya que a nivel central existe la capacidad de evaluar las respuestas a las órdenes dadas, tanto en circunstancias normales como en las sensorialmente conflictivas y en las patológicas.
Finalmente la información motora se organiza en dos sistemas de control que, formando parte de diferentes vías descendentes, actúan sobre la musculatura periférica:
- Sistema lateral (vía piramidal, vía rubrospinal), que actúa sobre la musculatura distal.
- Sistema medio (vía piramidal, vía vestibuloespinal, vía reticuloespinal), que actúa sobre la musculatura proximal y axial.
Este tipo de organización tiene como finalidad asegurar una tensión suficiente a los segmentos axiales y proximales para permitir el soporte de los segmentos distales y su orientación en el espacio pericorporal con vistas a tareas de prensión y manipulación. El sistema axioproximodistal está estrechamente coordinado con el sistema postural antigravitatorio, el primero tendría como soporte anatómico las vías reticuloespinales, mientras que el segundo ejercería su control a través de la vía vestibuloespinal; en ambos casos actuando sobre las motoneuronas, las interneuronas y las neuronas propioespinales.
El área premotora emite además eferencias directas sobre los núcleos neuromusculares del tronco del encéfalo y de la médula llevando a cabo una función independientemente de las zonas motoras primarias, interviniendo en la organización de los movimientos visuodirigidos y en las actividades vinculadas a la prensión de alimento y de la manipulación, características estas que se encontraban también en el neocerebelo
El área motora suplementaria está implicada en los movimientos autoiniciados en las tareas bimanuales y en las secuencias de actos motores. Sus vínculos con el área parietal 5 (área cinestésica), explican su probable función en la elección y estabilización de los segmentos corporales utilizados como referencia egocéntrico para el acto motor.
A pesar de la diversidad y complejidad de estos impulsos, el SNC consigue integrarlos en una única y significativa configuración para que el individuo tenga conciencia de su orientación espacial. Las diferentes áreas del córtex vestibular están organizadas jerárquicamente habiendo áreas de tipo primario y áreas de integración polisensorial desarrollando cada una aspectos particulares de la función vestibular.
Ahora bien, en el hombre todavía no se puede concretar mucho más sobre el córtex vestibular dada la ausencia de homología estricta entre las áreas corticales. Los conocimientos que se tienen al respecto proceden de la clínica, así crisis epilépticas con punto de partida parietal, temporal superior o insular, desencadenan sensaciones de rotación, vértigo, desequilibrio y distorsiones de las coordenadas visuales. Lo mismo se ha observado por estimulaciones peroperatorias en la fondo del surco intraparietal.
En el momento actual las investigaciones sobre la fisiología del equilibrio están desarrollando diversas técnicas experimentales para poder determinar la topografia las áreas del córtex vestibular basándose en su comportamieto a diversos estímulos vestibulares.
Las proyecciones vestibulocorticales son bilaterales con un predominio contralateral. Recordaremos que toman dos vías diferentes con una estación de interconexión en el tálamo, siendo la más importante la que se ubica en la parte lateral del núcleo verto-póstero-lateral.
Las proyecciones corticovestibulares son directas e indirectas y nacen del córtex parietal inferior.