- Generalidades.
- Anatomía del Sistema Nervioso Autónomo.
- Fisiología del Sistema Nervioso Autónomo.
- Simpático cefálico
- Simpático cervical.
 GENERALIDADES.
GENERALIDADES.
El Sistema Nervioso Autónomo constituye una parte del sistema nervioso central y periférico cuya función es el control y regulación de la homeostasis corporal.
Se denomina también Sistema Vegetativo y Sistema Órgano-Vegetativo. Este sistema, que funciona de forma inconsciente y autónoma, mantiene, controla y coordina el funcionamiento de los órganos y músculos a los que inerva, adaptando su funcionamiento en todo momento a las diferentes situaciones del medio ambiente externo e interno para mantener la situación de homeostasis del organismo. Entre otras funciones regula la respiración, la circulación, la digestión, el metabolismo, la secreción glandular, la temperatura corporal y la reproducción.
Estructura y división.
Por su anatomía y fisiológica el SNA se divide en tres partes: simpático, parasimpático y entérico.
- Simpático. Produce la movilización de las reservas corporales durante una emergencia o crisis emocional. Las pupilas se dilatan, la respiración se hace más profunda y aumenta el ritmo y la fuerza de las contracciones cardíacas. Los vasos sanguíneos de la piel y las vísceras, bajo su estímulo, se contraen, la presión sanguínea aumenta y hay mayor irrigación de los músculos esqueléticos, corazón, pulmones y cerebro.
- Parasimpático. Se relaciona principalmente con las actividades anabólicas relacionadas con la restauración y conservación de la energía corporal y el reposo de los órganos vitales. Así, estrecha la pupila para proteger a la retina de la luz excesiva, disminuye el ritmo cardíaco dando al músculo cardíaco periodos más prolongados de descanso. Es fundamental para la digestión de los nutrientes, ya que estimula la secreción salivar y los jugos gastrointestinales y estimula el peristaltismo digestivo.
- Entérico. Está formado por el el conjunto de los plexos submucoso de Meissner y mientérico de Auerbach, situados en la pared del tracto gastrointestinal. Está controlado por las divisiones simpática y parasimpática, si bien puede funcionar también por sí mismo en ausencia de inervación extrínseca. No vamos a tratar esta parte del SNA, pues su interés para el ORL es menor.
Los dos sistemas tienen un funcionamiento recíproco y sus actividades están integradas en respuestas coordinadas que aseguran el mantenimiento de un medio interno adecuado para afrontar las exigencias de una situación determinada.
ANATOMÍA DEL SISTEMA NERVIOSO AUTÓNOMO.
Cada parte del sistema autónomo, al igual que el sistema somático, consta de dos partes: una central y otra periférica.
Anatomía del Sistema Simpático.
 Centros superiores o centros diencefálicos.
Centros superiores o centros diencefálicos.
- El hipotálamo es el centro organizativo del SNA por excelencia, ya que controla todas las funciones vitales e integra el SNA con el sistema neuroendocrino. La áreas hipotalámicas relacionadas con el control del simpático son la ventromedial periventricular y la posterior. A destacar el núcleo posterolateral, de tal forma que un estímulo de esta zona genera una respuesta masiva de actividad simpática.
- Adhesión intertalámica: n. reuniens.
- Pared posterior del tercer ventrículo que está ocupada por la glándula pineal.
Estos centro presentan conexiones con la corteza cerebral, con la hipófisis, con los centros vegetativos de la región subtalámica y sublentiforme, con el mesencéfalo y con los centros del SNA situados por debajo de este nivel.
 Centros a nivel del troncoencéfalo.
Centros a nivel del troncoencéfalo.
- A nivel mesencefálico está el núcleo oculomotor accesorio que controla los movimientos de la pupila.
- En el rombencéfalo se encuentra los núcleos del SNA asociados a los de los pares craneales. Asociado al núcleo del facial están el núcleo salival superior y el núcleo lacrimomuconasal.
Centros a nivel medular.
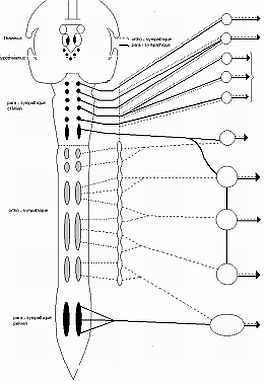
Estos centros autónomos vegetativos están situados en la sustancia gris medular, en la columna lateral (astas intermediolaterales), a lo largo de toda la región torácica. Los forman los cuerpos celulares de las primeras neuronas, o neuronas preganglionares de las vías simpáticas.
 Parte periférica.
Parte periférica.
Está formada por nervios y ganglios nerviosos.
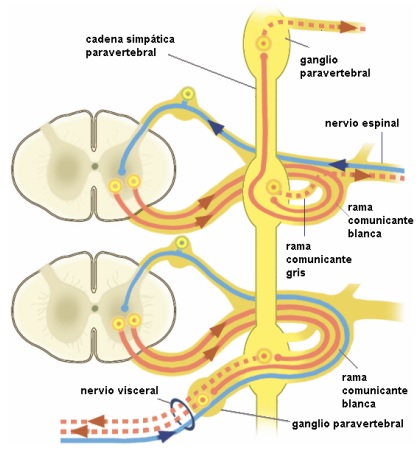
Los axones de las primeras neuronas situadas en la sustancia gris medular a nivel de los centros autónomos medulares se dirigen hacia el asta anterior de la médula para formar la raíz nerviosa ventral medular cuyos axones agrupados saldrán por el agujero de conjunción formando junto con la raíz dorsal, al juntarse ambas raíces, el nervio espinal o raquídeo mixto.
Estas fibras preganglionares simpáticas abandonan el nervio espinal inmediatamente después de la salida del agujero de conjunción a través de los nervioso denominados ramos comunicantes blancos. Estos ramos comunicantes que unen el nervio espinal con el ganglio simpático son de dos tipos: ramos comunicantes blancos cuyas fibras simpáticas son preganglionares comunicando los centros vegetativos medulares con los ganglios prevertebrales, y ramos comunicantes grises con un trayecto inverso uniendo los ganglios con los nervios espinales.
 Algunas fibras preganglionares no llegan al ganglio y forman el ramo meníngeo. Es un ramo nervioso destinado al contenido del conducto vertebral y a la columna vertebral. Está formado por dos raíces, una se desprende del nervio espinal antes de su división en sus dos ramos anterior y posterior; la otra procede del tronco simpático, directamente o mediante un ramo comunicante.
Algunas fibras preganglionares no llegan al ganglio y forman el ramo meníngeo. Es un ramo nervioso destinado al contenido del conducto vertebral y a la columna vertebral. Está formado por dos raíces, una se desprende del nervio espinal antes de su división en sus dos ramos anterior y posterior; la otra procede del tronco simpático, directamente o mediante un ramo comunicante.
Las fibras preganglionares son mielínicas delgadas de tipo B y amielínicas C.
Segunda neurona o neurona ganglionar: esta situada en los ganglios prevertebrales formando la cadena simpática.
Tronco o cadena simpática: está formada por dos troncos nerviosos situados en la cara anterior de la columna vertebral a la derecha y a la izquierda de la línea media uniendo una serie de ganglios escalonados. Recorre desde la base del cráneo hasta el cóccix. Los ganglios simpáticos se sitúan cerca de la médula espinal, para o prevertebrales, con lo que las fibras posganglionares tienen un trayecto relativamente largo.
Los ganglios son masas nerviosas de color grisáceo con variable volumen y forma, constituidas por los cuerpos de las segundas neuronas simpáticas, por ello los ganglios constituyen en si un centro nervioso vegetativo periférico que a su vez establece conexiones con: la médula espinal, los ganglios adyacentes y la periferia.
Cuando las fibras preganglionares entran en la cadena ganglionar, las fibras simpáticas pueden seguir tres caminos diferentes:
a) hacer sinapsis con las neuronas postganglionares del ganglio simpático del mismo nivel de la cadena simpática;
b) pueden dirigirse hacia arriba o hacia abajo y hacer sinapsis a otros niveles de la cadena;
c) pueden recorrer distancias variables dentro de la cadena simpática, y abandonarla sin hacer sinapsis. Estas fibras se dirigen hacia uno de los ganglios simpáticos colaterales, donde realizan sinapsis con la neurona postganglionar; estos ganglios son impares: ganglio celíaco, ganglio mesentérico superior y mesentérico inferior.
El recorrido más frecuentes es las fibras de las neuronas postganglionares localizadas en los ganglios simpáticos envien sus fibras desde estos al órgano efector. Sin embargo, un porcentaje de las fibras postganglionares simpáticas, viajan con los nervios somáticos (aproximadamente un 8% de las fibras de los nervios somáticos son simpáticas).
Comunicaciones y plexos nerviosos. Existen muchas comunicaciones mediante fascículos nerviosos entre los diferentes nervios. son particularmente abundantes en determinadas regiones donde forman un entrelazado de cordones y filetes nerviosos denominado plexo nervioso.
Vías simpáticas eferentes.
En es punto consideramos de interés hacer una aclaración histórica. En un principio, cuando se comenzó a tener un conocimiento del SNA, se mantuvo durante mucho tiempo como un concepto clásico que era un sistema puramente eferente- Esto fue debido a que el componente aferente no es tan claramente identificable. Hoy se conoce que el circuito nervioso de este sistema es muy similar, aunque con algunas importantes diferencias, al del resto de los circuitos del sistema somático: neuronas sensoriales de todo tipo y vías aferentes viscerales emiten señales que son conducidas a centro superiores de integración en el hipotálamo y el córtex cerebral y a órganos de la médula espinal, estas fibras aferentes constituyen el primer paso en los arcos reflejos. Estos órganos nerviosos son los encargados de elaborar una respuesta que es transportada mediante vías eferentes viscerales y de tipo motor, simpáticas y parasimpáticas a los efectores: músculos y vísceras secretoras. Las respuestas pueden ser de dos tipos: contracción de músculos involuntarios o secreción glandular.
Las vías simpáticas eferentes nacidas de la segunda neurona pueden ser somáticas o viscerales. Las fibras postganglionares son mayoritariamente amielínicas.
Fibras somáticas: Se originan del ganglio del tronco simpático y desde aquí alcanzan el nervio espinal mediante un ramo comunicante gris para aportar a este su componente vegetativo y se dirigen a inervar el mismo territorio que los nervios espinales: tejidos y órganos periféricos.
Fibras viscerales: el ganglio en que se origina la segunda neurona es un ganglio periférico que se halla próximo o distante de las vísceras, para desde allí dirigirse al órgano al que están destinadas a inervar ya sea directamente o después de conformar un plexo nervioso simpático.
Vías simpáticas aferentes.
Conducen la sensibilidad visceral y vascular mediante dos vías:
Vías cortas: llevan la sensibilidad de la víscera al ganglio periférico y desde el cuerpo celular se envía hacia otros centros y vías motoras.
Vías largas: van al tronco simpático, al nervio espinal por el ramo comunicante blanco y alcanzan su cuerpo celular en el ganglio espinal de la raíz dorsal, este axón alcanza las diversas vías de la sensibilidad general consiente o inconsciente.
Anatomía del Sistema Parasimpático.
Este sistema consta de dos porciones una craneana y otra pélvica.
Porción Parasimpática Craneal.
Centros superiores o centros diencefálicos: a nivel hipotalámico el parasimpático es controlado por las áreas periventricular lateral y anterior conocidas como núcleos hipotalámicos medial y anterior.
Porción Parasimpática troncoencefálica.
Las fibras nacidas a partir de centros superiores mesencefálicos alcanzan los núcleos de nervios craneanos y parten hacia el organismo a partir se sus nervios: oculomotor, facial, intermedio, glosofaríngeo y vago.
Nervio Oculomotor (III): es portador de las fibras parasimpáticas que nacen de núcleo accesorio del nervio oculomotor y por medio de la raíz parasimpática estas fibras se dirigen al ganglio ciliar del que emerge la fibra posganglionar formando ramos ciliares cortos, que dan inervación al músculo ciliar y al esfínter de la pupila.
Nervio Facial (VII): sus fibras parasimpáticas parten del núcleo lagrimal, discurriendo a través del nervio petroso mayor se dirigen al ganglio pterigopalatino, y desde éste por medio del nervio cigomático y maxilar terminan en la glándula lagrimal; y por medio de los ramos nasales posteriores, superiores palatinos menores y faríngeos, se dirigen a la mucosa nasal.
Nervio Intermediaro: parten fibras parasimpáticas del núcleo salivatorio superior, las cuales se dirigen al tronco del facial, continúan a los ganglios submandibular y sublingual, desde aquí las fibras pos ganglionares se dirigen a la glándula correspondiente.
Nervio Glosofaringeo (IX): sus fibras parasimpáticas nacen del núcleo salivatorio inferior que constituyen el nervio petroso menor que pasa a través del tímpano y termina en el ganglio ótico, desde aquí las fibras posganglionares se dirigen por medio del nervio auriculotemporal hasta la parótida.
Nervio Vago (X): sus fibras preganglionares parten del núcleo dorsal del vago, desde aquí algunas alcanzan diversos plexos simpáticos y hacen sinapsis en sus ganglios. Aproximadamente un 75% de las fibras del SNP viajan a través del nervio vago hacía las regiones torácica y abdominal. El vago, proporciona fibras parasimpáticas al corazón, pulmones, esófago, estómago, intestino delgado, mitad proximal del colon hasta la flexura esplénica, hígado, vesícula.
Porción parasimpática pélvica.
Las fibras preganglionares se originan en centros ubicados en la sustancia gris intermediolateral de la porción sacra de la médula espinal, desde donde salen como ramos comunicantes blancos, se incorporan a las ramas anteriores de los nervios sacros S2, S3 y S4, formando nervios esplácnicos pelvianos que se unen al plexo hipogástrico. Desde aquí las fibras posganglionares ocupan su territorio en la parte terminal del intestino grueso, la vejiga, órganos genitales internos y órganos eréctiles.
Los ganglios parasimpáticos yacen cerca o dentro de las estructuras viscerales, por lo que sus fibras postganglionares son cortas. Todos los nervios simpáticos y parasimpáticos incluyen fibras eferentes viscerales, sin embargo, los nervios parasimpáticos tienen de promedio el triple de axones sensitivos que sus homólogos simpáticos. La mayoría de estas fibras eferentes viscerales son amielínicas.
El simpático y el parasimpático funcionan en general de forma antagónica y el efecto final depende del balance entre los dos, esto supone que los diferentes órganos han de recibir inervación de ambos sistemas. Sin embargo, hay órganos que solo reciben inervación simpática o parasimpática.
Diferencias entre el sistema somático y el sistema autónomo.
- La mayor diferencias entre ambos sistemas nerviosos es que el sistema nervioso autónomo no se encuentra sujeto al control voluntario o consciente. La mayoría de las señales sensoriales viscerales que recibe el SNC no son percibidas de manera consciente. La función de estas señales es la de efectuar ciertos cambios en el comportamiento dirigidos a responder de la forma más adecuada al estímulo.
- La cadena neural eferente de ambos sistemas es diferente. Los efectores somáticos están inervados por una sola neurona, mientras que la cadena efectora de los dos sistemas autónomos consta de dos neuronas. Las neuronas somáticas se proyectan directamente sobre sus efectores, sin embargo las neuronas del sistema autónomo emiten sus eferencias a través de una cadena formada por dos neuronas: una neurona preganglionar, situada en en el sistema nervioso central medular, cuyo axón va a sinaptar con la segunda neurona que es la ganglionar, cuyo soma está en un ganglio nervioso autonómico y que será la que inerve los órganos diana. Luego hablaremos de los ganglios nerviosos del sistema autónomo donde se encuentran las cc efectoras.
- Las neuronas motoras somáticas activan únicamente fibras musculares esqueléticas, mientras que las neuronas motoras autónomas tienen actuación sobre una mayor diversidad de blancos: músculos lisos, músculo cardíaco, endotelios y glándulas de secreción.
- Las comunicaciones entre los nervios del sistema somático se efectúa entre dos nervios vecinos de la misma mitad del cuerpo y entre nervios que comparten el mismo territorio de inervación. Los plexos de este sistema se forma sobre todo en los troncos nerviosos que se dirigen a los miembros, p.e. el plexo braquial. Por el contrario las comunicaciones entre los nervios del sistema vegetativo se realizan entre los nervios de un lado con los del lado contrario, además los plexo que se forma son sumamente complejos y a través de ellos el nervio de un lado participa en la inervación de ambos lados de las vísceras que va a inervar.
- En su distribución el SNA sigue el denominado principio de divergencia del estímulo: cada neurona preganglionar hace sinapsis con un gran número de neuronas posganglionares, permitiendo así que un pequeño número de neuronas centrales influyan sobre grandes áreas corporales. Una neurona preganglionar puede hacer sinapsis posganglionares tanto excitadoras como inhibidoras, permitiendo la inhibición o excitación de numerosas cc blanco, lográndose un mayor grado de coordinación funcional.
- En cuanto a su fisiología los ganglios nerviosos del SNA tienen un cierto grado de autonomía funcional con respecto al SNC ya que los axones eferentes que transcurren por los ganglios autónomos proporcionan a los ganglios una gran parte de la integración sensorio-motora.
FISIOLOGÍA DEL SISTEMA NERVISO AUTÓNOMO.
La integración de las actividades del sistema nervioso autónomo ocurre a todos los niveles del eje cerebroespinal. El SNC, como centro superior, integra continuamente los dos sistemas efectores, somático y autonómico, ambas partes del sistema nervioso no funcionan de manera aislada ni completamente independiente. Las órdenes que se originan en el cerebro y las respuestas reflejas desencadenadas por estímulos generales o específicos siguen dos vías efectoras para controlar las respuestas de adaptación y de relación, que actúan sobre los músculos esqueléticos y sobre los órganos viscerales. Además, el SNA está modulado y refleja las variaciones de la actividad subjetiva y emocional.
La corteza cerebral es el nivel más alto de integración somática del SNA. Fruto de esta integración, un único estímulo puede desencadenar respuestas autonómicas, antinociceptivas y de comportamiento.
La activación del SNA se produce fundamentalmente a partir de centros localizados en el hipotálamo. El hipotálamo es el centro organizativo por excelencia, ya que controla todas las funciones vitales e integra el SNA con el sistema neuroendocrino.
Los estímulos u ordenes centrales son derivadas a estaciones nerviosas intermedias situadas en el troncoencéfalo donde se localizan los centros de la respuesta aguda del SNA, así p.e. en el bulbo encefálico se encuentra el principal centro de integración de los reflejos cardiovasculares.
Finalmente la parte periférica se sitúa en los centros autónomos la medula espinal que son el sustrato de los reflejos viscerales. La mayor parte de las funciones del SNA son reguladas de forma inconsciente, mediante reflejos viscerales, aunque pueden ser influenciadas por las emociones y los estímulos somatosensoriales.
 Neurotransmisión del SNA.
Neurotransmisión del SNA.
Los neurotransmisores del sistema nervioso simpático y parasimpático son fundamentalmente la noradrenalina (NA) y la acetilcolina (AC). Las fibras secretoras de NA se denominan adrenérgicas y las que secretan AC, colinérgicas.
Neuronas preganglionares: tanto las del sistema nervioso simpático como las del parasimpático, son colinérgicas.
Neuronas postganglionares: los neurotransmisores secretados por la neurona postganglionar van a reaccionar con los receptores de los órganos a los que inervan provocando la respuesta biológica. Las neuronas postganglionares del sistema nervioso simpático son adrenérgicas y secretan noradrenalina con excepción hecha de las fibras que van a las glándulas sudoríparas y a una minoría de vasos sanguíneos que son colinérgicas. Las que se dirigen a la médula suprarrenal secretan sobre todo adrenalina y muy poca cantidad de noradrenalina. Las neuronas postganglionares del parasimpático son colinérgicas.
Además de la noradrenalina y de la acetilcolina, existen otros neurotransmisores que también están relacionados con la transmisión nerviosa en el sistema nervioso autónomo aunque tengan menos importancia: Óxido nítrico, Péptido intestinal vasoactivo, Adenosina Trifosfato, Neuropéptido Y, Serotonina, Ácido gamma‐amino butírico, Hormona liberadora de gonadotropina y Sustancia P. La Dopamina también se considera un neurotransmisor adrenérgico sin serlo, ya que es un precursor en la síntesis de noradrenalina y adrenalina.
La liberación de neurotransmisores en la hendidura sináptica a partir de las vesículas intracelulares, ocurre por un proceso de exocitosis. Los neurotransmisores difunden a través de la hendidura sináptica uniéndose a los receptores postsinápticos.
El SNA realiza un control multisistémico de forma continua y constante a lo largo de los ciclos vitales, tanto en reposo como en actividad. En situación de reposo se liberan continuamente pequeñas cantidades o "cuantos" de neurotransmisor, cada uno de los cuales origina pequeños cambios eléctricos, "potenciales en miniatura", en la membrana postsináptica que no llegan a desencadenar la despolarización. La llegada de un potencial de acción provoca la liberación sincrónica de cientos de cuantos, que sí causa la despolarización de la membrana postsináptica.
La liberación de los neurotransmisores está influenciada por múltiples factores, pero hay dos mecanismos básicos:
- Interacción heterotrófica: no se liberan por estimulación directa de la terminación liberadora sino bajo la influencia de otras neuronas cercanas, por ej. neuronas simpáticas pueden inhibir la actividad de neuronas parasimpáticas cercanas y viceversa.
- Interacción homotrópica o “feed‐back” auto‐inhibitorio: la membrana presináptica puede tener receptores para sus propios neurotransmisores, que son estimulados simultáneamente a los postsinápticos, inhibiendo la liberación de más neurotransmisores.
Las terminaciones postganglionares del sistema nervioso simpático y parasimpático son anatómica y fisiológicamente similares, sólo las diferencia el neurotransmisor; presentan múltiples ramificaciones que constituyen, como hemos dicho, los plexos efectores terminales que envuelven o rozan la unidad efectora. La mayor parte de las vísceras reciben una inervación doble del SNA, simpática y parasimpática. En general, las dos divisiones funcionan integradamente en el control visceral y ejercen efectos opuestos sobre cada órgano. Ahora bien, algunas estructuras viscerales, como las glándulas sudoríparas o la mayoría de los vasos sanguíneos, reciben inervación solamente de uno de los dos sistemas.
Síntesis y receptores de los neurotransmisores.
La velocidad de síntesis de neurotransmisor depende de las necesidades, y por ende, del grado de actividad del SNA, y está regulada por mecanismos de “feed‐back” local.
 Adrenalina y noradrenalina.
Adrenalina y noradrenalina.
La síntesis de noradrenalina se inicia en el axoplasma de las fibras adrenérgicas y acaba almacençándose en las vesículas presinápticas de las terminaciones nerviosas. En el axoplasma se sintetiza la dopamina y a continuación ésta es transportada hasta el interior de las vesículas donde se formará la noradrenalina. En la médula suprarrenal se lleva a cabo una última etapa para la formación de adrenalina. El paso limitante en la síntesis de noradrenalina es el paso de tirosina a Dopa y aquí se controla la producción mediante un mecanismo de “feed‐back” negativo.
La eliminación de la noradrenalina, una vez secretada en la sinapsis ocurre de las siguientes formas:
- El 50‐80% de la noradrenalina es recuperada hacia el interior de las terminaciones adrenérgicas presinápticas por transporte activo para ser reutilizada, siendo este el principal mecanismo de finalización de la acción de la noradrenalina. Este proceso de recaptación se realiza por proteínas transportadoras específicas localizadas en la membrana presináptica. Una gran variedad de factores puede influenciar y alterar la actividad del transportador como la exposición al etanol, ciertos fármacos, activación o inhibición de receptores presinápticos, etc.
- La noradrenalina restante difunde hacia los fluidos corporales vecinos y de aquí a la sangre sufriendo metabolización hepática y renal; esta última es la principal vía metabólica de las catecolaminas administradas exógenamente.
- Una pequeña cantidad de noradrenalina es metabolizada, por la monoaminoxidasa en las propias terminaciones nerviosas o por la catecolmetiltransferasa presente en el resto de los tejidos, formándose ácido vanil‐mandélico que se eliminará por la orina.
- Menos de un 5% de la noradrenalina se elimina de forma inalterada por la orina.
Para estos neurotransmisores existen 2 grande grupos de receptores adrenérgicos, los alfa (α) y los beta (β), que a vez se subdividen en alfa1, alfa2, beta 1 y beta2, aunque se conocen más subdivisiones. A “grosso modo” los alfa1 serían los postsinápticos, los alfa2 presinápticos, los beta1 cardíacos y los beta2 no cardíacos.
Los adrenorreceptores se localizan tanto en la neurona presináptica como en la postsináptica, así como también en localizaciones extrasinápticas. Se localizan tanto a nivel post como presináptico, y también pueden ser extrasinápticos. Los receptores alfa se estimulan sobre todo por la acción de la noradrenalina, mientras que los receptores beta se estimulan sobre todo con el isoproterenol. Existe además, otro receptor periférico adrenérgico específico para la dopamina, el receptor dopaminérgico.
Los receptores presinápticos se consideran inervados ya que se encuentran muy próximos a la liberación del neurotransmisor. Los postsinápticos pueden considerarse inervados o no dependiendo de su proximidad a la hendidura sináptica, de manera que los que están en la misma membrana postsináptica se consideran inervados y los extrasinápticos se denominan no inervados.
Acetilcolina.
La síntesis de acetilcolina tiene lugar en la terminación presináptica de las fibras colinérgicas mediante la acetilación de la colina con acetil‐coenzima A. Esta reacción esta catalizada por la acetilcolintransferasa. La mayor parte de la síntesis ocurre en el axoplasma y posteriormente es transportada a las vesículas sinápticas donde queda almacenada. Una vez liberada en la sinapsis, permanece en la sinapsis durante pocos segundos, tiempo durante el cual se produce la respuesta biológica.
A continuación, se elimina de forma muy rápida gracias a la hidrólisis por la acetilcolinesterasa, enzima presente en la neurona, en la hendidura sináptica y en diversos tejidos, dando lugar a iones acetato y a colina. La colina es reutilizada, siendo transportada hacia el interior de la neurona presináptica mediante transporte activo, para la síntesis de más acetilcolina. Una mínima cantidad de acetilcolina difunde hacia líquidos vecinos y plasma donde se metabolizará por la pseudocolinesterasa o colinesterasa plasmática, pero esta enzima tiene muy poca importancia en la metabolización de la acetilcolina. La acetilcolina como tal no se reutiliza y debe sintetizarse de forma constante.
La acetilcolina es el neurotransmisor del sistema nervioso parasimpático, de ganglios del sistema nervioso simpático y en la unión neuromuscular estriada. Los receptores sobre los que actúa la acetilcolina, se conocen como colinérgicos. Los receptores colinérgicos, se diferencian en dos tipos: los muscarínicos y los nicotínicos, según su afinidad selectiva para la muscarina o la nicotina.
Los receptores muscarínicos pertenecen al grupo de los receptores metabotrópicos que usan proteínas G como mecanismo para dar señal. Se encuentran en los órganos efectores estimulados por las neuronas postganglionares del sistema nervioso parasimpático, como el corazón y el músculo liso de todo el organismo. Existen cinco tipos de estos receptores. La estimulación de los receptores muscarínicos produce bradicardia, disminución del inotropismo, broncoconstricción, miosis, salivación, hipermotilidad gastrointestinal y aumento de la secreción de ácido gástrico. Estos receptores pueden bloquearse con atropina sin que se produzcan efectos sobre los receptores nicotínicos.
Los receptores nicotínicos se localizan en las uniones sinápticas de las neuronas pre y postganglionares tanto del simpático (ganglios simpáticos) como del parasimpático; así el estímulo nicotínico produce efectos excitatorios en ambos sistemas, pero el efecto final es predominantemente simpático con hipertensión y taquicardia a través de la liberación de adrenalina y noradrenalina de la médula suprarrenal. Los receptores de la unión neuromuscular estriada son también nicotínicos, pero de un tipo diferente a los autonómicos.
Funcionamiento interactivo de ambos sistemas.
Ambos sistemas eferentes ejercen un control de la función de los órganos viscerales ya que estos están inervados simultáneamente por fibras nerviosas simpáticas y parasimpáticas.
Sin embargo algunos órganos efectores reciben inervación de un solo sistema. Por ejemplo, el músculo liso de la mayoría de los vasos sanguíneos está controlado únicamente por la inervación simpática. En estos casos, el control de la actividad visceral depende de las variaciones de la frecuencia de descarga de los impulsos por la inervación simpática.
Los nervios autonómicos presentan una actividad tónica basal (de frecuencias de impulsos bajas, de 0,5 a 5 impulsos por segundo), partiendo de este tono basal esta actividad puede aumentar o disminuir. En los vasos, un aumento de frecuencia simpática determina vasoconstricción y una disminución vasodilatación. La actividad del órgano depende de la interacción o equilibrio entre las señales de ambos sistemas, que ejercen efectos antagónicos. Esta interacción de ambos sistemas puede desarrollarse por acciones contrarias sobre las mismas cc efectoras, como ocurre en el corazón, donde el simpático excita las células nodales al aumentar la frecuencia cardíaca, mientras que el parasimpático inhibe las mismas células al reducir la frecuencia. Otra posibilidad es la actuación sobre cc distintas, de efectos contrarios; así, en el iris, el simpático excita las fibras musculares meridianas, lo que produce midriasis, mientras que el parasimpático excita las fibras circulares y produce miosis.
En circunstancias normales, los dos sistemas son recíprocamente activos, debido a que las señales centrales y reflejas excitan un sistema e inhiben el otro. Cuando ambos sistemas, simpático y parasimpático, inervan cc dianas próximas se registran interacciones más complejas; por ejemplo, la activación simultánea de las fibras simpáticas puede exagerar la respuesta cardiovascular a la actividad parasimpática y viceversa.
Una de sus principales características del funcionamiento del SNA es la rapidez y la intensidad con la que puede cambiar las funciones viscerales. Así por ejemplo, en cuestión de 3‐5 segundos puede duplicar la frecuencia cardiaca y en 10‐15 segundos la presión arterial. No obstante el SNA está diseñado para producir acciones más bien lentas y sostenidas, en comparación con las del sistema nervioso somático.
Reflejos autónomos.
Muchas de las funciones viscerales del cuerpo están reguladas por circuitos reflejos autonómicos. Los componentes eferentes del SNA se activan o inhiben por las señales aferentes de vías reflejas, independientes de la voluntad. Sin embargo, estos circuitos reflejos son frecuentemente modulados por las señales de estructuras centrales que integran las respuestas somáticas y autonómicas y por eso en actos reflejos fundamentalmente somáticos existen respuestas viscerales acompañantes; por ejemplo, ante un estímulo doloroso de cierta intensidad, se produce una respuesta motora en forma de reflejo de retirada, pero concomitantemente aparecen respuestas autonómicas, como taquicardia y aumento de la presión arterial. De la misma manera, los reflejos fundamentalmente viscerales suelen ir acompañados de reacciones somáticas; así, ante un dolor intestinal intenso se produce una respuesta refleja autonómica, que tiende a paralizar la actividad intestinal; en paralelo, se puede apreciar una respuesta somática de contracción muscular abdominal (que puede ser bastante intensa dando un ‘abdomen en tabla’).
Los circuitos reflejos autonómicos en general son muy complejos, entre ellos las conexiones polisinápticas entre vías aferentes y eferentes a varios niveles del SNC.
Los reflejos autónomos más importantes son
- Reflejo barorreceptor que mediante el control del calibre de los vasos sanguíneos controla la tensión arterial
- Reflejos pupilares que controla el diámetro de la pupila.
- Control de la temperatura corporal.
- Reflejo de la micción.
- Respuestas sexuales.
Salvo algunas excepciones el dolor visceral está mediado por axones aferentes de los nervios simpáticos. En tanto que la información proveniente de las víscera necesaria para su regulación está mediada por axones aferentes que transcurren por nervios parasimpáticos.
Las disfunciones del SNA comportan manifestaciones clínicasmúltiples y variables, que, a menudo, son infravaloradas en la prác-tica clínica. Sin embargo, los síntomas o alteraciones autonómicosdeben tenerse siempre en cuenta por su implicación diagnóstica,por la repercusión sobre la calidad de vida del paciente y por lainfluencia pronóstica sobre la expectativa de vida.
La información sensitiva que llega al SNA tiene tres orígenes: vísceras, sensibilidad visceral, superficie corporal, sensibilidad somática y el medio exterior, sensibilidad especial.
SISTEMA SIMPÁTICO CEFÁLICO.
El tronco cervical simpático, o cadena simpática cervical, es una continuación cefálica del tronco simpático torácico. Se encuentra en un espacio fasciculado y limitado; por detrás por la fascia de los músculos paravertebrales y por delante por la vaina de la carótida El simpático cefálico está formado por tres ganglios principales unidos al simpático cervical por el plexo carotídeo y por la arteria meníngea media. Son los ganglios oftálmico, esfenopalatino y ótico.
Cada uno de estos tres ganglios posee ramas aferentes, periféricas o eferentes y anastomóticas.
 Ganglio oftálmico.
Ganglio oftálmico.
Es pequeño, cuadrilátero, alargado de adelante a atrás. Aplanado transversalmente y situado en la cara externa del nervio óptico, a siete u ocho milímetros por delante del agujero óptico.
Como ramos aferentes recibe por su extremidad posterior tres raíces.
1) Una raíz motora que va del nervio del oblicuo menor, rama del MOC, al ángulo póstero inferior del ganglio.
2) Una raíz sensitiva que une el nervio nasal al ángulo póstero-superior del ganglio.
3)Una raíz simpática que nace del plexo carotídeo y termina en el borde posterior del ganglio oftálmico entre las dos raíces anteriores.
Los ramos eferentes nacen del borde anterior del ganglio oftálmico, denominados nervios ciliares cortos, en número de cinco a seis. Se dirigen hacia delante, dividiéndose y perforando la esclerótica alrededor del nervio óptico. Están destinados a la esclerótica de la cornea, a la coroides, al cuerpo ciliar y al iris.
Hay un ramito nervioso que se desprende de los nervios ciliares por detrás del globo ocular que se le denomina nervio de Tiedemann y que penetra, junto con la arteria central de la retina, en el nervio óptico.
Ganglio esfenopalatino.
Es un abultamiento que se encuentra en la extremidad anterior del nervio vidiano. Se halla en la pared posterior del transfondo de la fosa ptérigo-maxilar, en la fosita en forma de embudo que continúa el conducto vidiano. El ganglio, de forma cónica, está unido al nervio vidiano por su vértice dirigido hacia atrás.
El ganglio esfeno-palatino recibe al nervio vidiano. Este nervio está formado por la reunión del nervio petroso superficial mayor, rama del NF, del nervio petroso profundo mayor, rama del glosofaríngeo y de un ramo simpático procedente del plexo pericarotídeo. Desde su emergencia en el hiato de Falopio los nervios petrosos mayores superficial y profundo circulan de atrás a delante por la cara ántero-superior del peñasco, cruzan la cara inferior del ganglio de Gasser o la cara inferior del nervio maxilar inferior y se unen. El tronco nervioso que resulta recibe la raíz simpática unas veces por encima de lámina fibrosa que obtura el agujero rasgado anterior y otras veces en su espesor.
Los ramos eferentes del ganglio esfeno-palatino terminan en el nervio esfeno-palatino que desciende por fuera o por delante de él y se le adhiere. Estos ramos terminan en la mucosa buco-naso-faríngea. Otras fibras del ganglio, en conexión con el facial por el nervio petroso superficial mayor, se dirigen indirectamente por medio del nervio maxilar superior al ramo orbitario de este nervio y por el nervio lagrimal, con el cual este último se anastomosa, a la glándula lagrimal.
Ganglio ótico.
Es un ganglio minúsculo de dos o tres mm de diámetro. Está situado por debajo del agujero oval, en la cara interna del nervio maxilar inferior o de la extremidad superior del nervio lingual.
En este ramo termina un ramo nervioso formado por la reunión del nervio petroso superficial menor, del nervio petroso profundo menor y de un ramo simpático del plexo que rodea la arteria meníngea media. Los ramos que parten del ganglio ótico van a las ramas sensitivas del maxilar inferior, en particular al nervio aurículo-temporal. Estos últimos inervan por medio de él, la glándula parótida. Algunas fibras van también al tronco común de los nervios del periestafilino externo, del pterigoideo interno, del músculo del martillo y a la cuerda del tímpano.
Ganglio submaxilar.
Situado entre el nervio lingual y el borde superior de la glándula submaxilar. Recibe conexiones del nervio lingual, de la cuerda del tímpano y del plexo simpático de la arteria facial. Emite ramos destinados a la glándula submaxilar.
Ganglio sublingual.
Está en el trayecto del nervio sublingual, en la parte posterior o en la cara externa de la glándula sublingual.
Como el submaxilar, recibe conexiones del nervio lingual, de la cuerda del tímpano y del plexo simpático de la arteria facial. Emite ramos destinados a la glándula sublingual.
SISTEMA SIMPATICO CERVICAL
Comprende dos o tres ganglios unidos por un cordón intermedio conocidos como cadena simpática cervical. Constituye el centro cérvico-mediastínico anterior, que contribuye a la formación en el tórax de los plexos cardíaco y pulmonar. Está situado detrás del paquete vásculo nervioso y aplicado a la aponeurosis prevertebral por una delgada lámina célulo-fibrosa, que puede ser considerada como una hoja de desdoblamiento de esta aponeurosis. La aponeurosis y los músculos prevertebrales separan el simpático de las apófisis transversas de las vértebras cervicales.
Ganglios cervicales.
Son tres: superior, medio e inferior.
El ganglio cervical superior es voluminoso y fusiforme. Su longitud es de unos cuatro cm. Por detrás está en relación con el recto anterior y con la segunda y tercera vértebras cervicales. Por delante se relaciona con el paquete vásculo-nervioso retroestiloideo del espacio máxilo-faríngeo.
El ganglio cervical medio es inconstante. Cuando existe, es minúsculo. Está situado entre la apófisis transversa de la sexta vértebra cervical por detrás y el paquete vásculo-nervioso por delante.
El ganglio cervical inferior o ganglio estrellado. Se encuentra situado en la cadena simpática cervical entre el cervical medio y el primer ganglio torácico. Es un poco aplanado de delante a atrás y alargado de arriba abajo. Su longitud es de 8 mm aproximadamente. Su forma es muy variable, unas veces redondeado, otras semilunar, y otras estrellado.
Está situado entre el cuello de la costilla por delante y la apófisis transversa de la séptima vértebra cervical por detrás. En algunas ocasiones puede estar en situación más baja descendiendo por detrás de la pleura y por delante del primer espacio intercostal, hasta la primera costilla. Esta situado por encima y por detrás de la cúpula pleural, en una depresión denominada fosita supra-retro pleural, limitada por el ligamento vértebra-pleuro-costal, por encima y por fuera, y por último, por la extremidad posterior de la primera costilla, por detrás. La arteria y la vena vertebrales pasan por delante de él y lo cubren, pero, a menudo, el ganglio desborda la arteria hacia dentro, o más frecuentemente hacia fuera. La arteria subclavia está separada de él por la parte más alta de la cúpula pleural. El tronco cérvico-intercostal cruza su cara externa.
En la mayoría de los casos este ganglio cervical se compone dos porciones, unidas entre sí por cordones o filetes nerviosos más o menos numerosos. Una de estas porciones ocupa el lugar del ganglio normal, la otra, denominada ganglio intermedio, está situada por encima y por dentro de la precedente sobre el lado interno de la arteria vertebral. Existe habitualmente una relación inversa entre el tamaño de estas dos masas ganglionares. Así cuando una de ellas es pequeña la otra es más voluminosa, y viceversa.
Cordón intermedio.
Los tres ganglios cervicales están unidos entre sí y a la cadena torácica por el cordón intermedio. A nivel del ganglio medio, el cordón se desdobla algunas veces en dos ramos, que abrazan el primer codo de la arteria tiroidea inferior. Se divide después en dos cordones secundarios. Uno continúa el trayecto del cordón principal y va al ganglio cervical inferior. Otro se compone habitualmente de varios filetes nerviosos que se desprenden unas veces del ganglio cervical medio, otras del cordón intermedio por debajo de este ganglio, y otras, que es lo más frecuente, del ganglio estrellado. Estos filetes descienden por delante de la arteria vertebral, pasan después sucesivamente, por las caras anterior, inferior y posterior de la arteria subclavia, y terminan en el ganglio estrellado. De este modo describen alrededor de la arteria subclavia una curva cóncava hacia arriba y hacia atrás, que se conoce con el nombre de asa de Vieussens.
Ramas anastomóticas de los ganglios cervicales.
El ganglio cervical superior está anastomosado con:
-
Los ganglios yugular y de Andersch, por un ramo llamado nervio yugular.
-
Directamente con el glosofaríngeo, neumogástrico, hipogloso mayor y las ramas anteriores de los tres o cuatro primeros nervios cervicales.
-
El ganglio cervical superior, mediante el denominado nervio carotídeo, está unido a los ganglios esfeno-palatino y oftálmico, así como a los nervios que caminan por la pared del seno cavernoso y al ganglio de Gasser. Este nervio parte del polo superior del ganglio, asciende por detrás de la carótida interna, penetra con ella en el conducto carotídeo y forma a su alrededor un rico plexo denominado plexo cavernoso de que se van desprendiendo todos estos filetes anastomóticos. El plexo cavernoso contiene además algunos ganglios simpáticos.
-
El ganglio cervical medio está unido por ramos comunicantes con las ramas anteriores de los nervios cervicales quinto y sexto.
-
El ganglio cervical inferior está unido directamente a los séptimo y octavo nervios cervicales, al primero dorsal, al nervio cardiaco medio y al frénico por ramos distintos. Además está unido por el nervio vertebral al cuarto, quinto y sexto nervios cervicales.
Ramas periféricas de los ganglios cervicales.
-
Ramas del ganglio cervical superior. Da ramos vasculares, faríngeos, esofágicos, laríngeos y el nervio cardíaco superior.
Los ramos vasculares o carotídeos, en número variable, descienden a lo largo de la carótida interna. Se anastomosan con ramos del vago y del glosofaríngeo y forman con ellos, a nivel de la bifurcación de la carótida primitiva, el plexo intercarotídeo. De este plexo se desprenden numerosos ramitos que constituyen plexos secundarios alrededor de las diferentes ramas colaterales de la carótida externa. El plexo intercarotídeo inerva también el seno carotídeo y la glándula intercarotídea.
Los ramos faríngeos, numerosos, forman el plexo faríngeo, anastomosándose en la pared lateral de la faringe con ramos del glosofaríngeo y del neumogástrico
Ramos esofágicos que van a la parte superior del esófago.
Ramos laríngeos que se anastomosan con ramos de los nervios laríngeos del neumogástrico para formar, por dentro de la carótida primitiva, el plexo laríngeo de Haller. De este plexo salen ramitos destinados a la laringe, tiroides y esófago.
El nervio cardíaco superior nace también de este ganglio.
-
Ramos del ganglio cervical medio. Son ramos vasculares que acompañan a la arteria tiroidea inferior y además al nervio cadíaco medio, que termina en el plexo cardíaco posterior.
-
Ramos del ganglio cervical inferior. Da ramos vasculares, el nervio cardíaco inferior y algunos ramitos que van a lo largo del cuello o a la primera vértebra dorsal.
Los ramos vasculares están destinados a la arteria subclavia y a sus ramas. Uno de ellos, voluminoso, satélite de la arteria vertebral, es conocido con el nombre de nervio vertebral.
El nervio vertebral nace de este ganglio por dos clases de raíces. Unas anteriores, ascienden por delante de la arteria y se pierden alrededor de ella. Otras raíces, generalmente en número de dos, son posteriores con respecto a la arteria. Unas y otras, se reúnen en un tronco común que es el nervio vertebral, el cual asciende hasta la cuarta vértebra cervical, por detrás de la arteria vertebral que, a su vez, se encuentra por dentro de la vena. Aplicado sobre la arteria, el nervio está situado enfrente del intersticio que separa ambos vasos. Da, al pasar, un ramo comunicante para los nervios cervicales séptimo, sexto y quinto y algunas veces también para el cuarto. El ramo comunicante destinado al cuarto cervical parece ser el terminal del nervio vertebral. Este ramo comunicante da ramos vasculares delgados que rodean la arteria formando un plexo que se prolonga hasta el interior de la cavidad craneal, alrededor del tronco basilar.
El nervio vertebral da origen, además, a la raíz simpática de los nervios sinuvertebrales correspondientes.
El nervio cardíaco inferior contribuye a formar el plexo cardíaco posterior.
Los nervios cardíacos superior, medio e inferior representan los esplácnicos mediastínicos anteriores.
La mayor parte de las funciones del SNA son reguladas de forma inconsciente, mediante reflejos viscerales, aunque pueden ser influenciadas por las emociones y los estímulos somatosensoriales. La corteza cerebral es el nivel más alto de integración somática del SNA
Para seguir visualizando este capítulo descargar tema completo![]()