- FUNCIONES.
- ESTÍMULOS DE LOS RECEPTORES VESTICULARES.
- VESTIBULARES.
- FISIOLOGÍA DE LOS RECEPTORES MACULARES.
- FISIOLOGÍA DE LOS RECEPTORES CANALICULARES.
- FISIOLOGÍA DEL NEUROEPITELIO.
- NERVIO VESTIBULAR.
- NÚCLEOS VESTIBULARES.
- CONEXIONES INTERVESTIBULARES.
El SV es un elemento tan específico del sistema del equilibrio que se le ha denominado "órgano del equilibrio", pero esta denominación tiene tan sólo un fondo de admiración, más que una base firme y real.
Cave destacar que de los tres receptores de la tríada sensorial del equilibrio, el SV es el receptor de información más específico de la función equilibrio, ya que si bien los tres contribuyen a esta función, los otros dos receptores, visual y propioceptivo, tienen además otras funciones.
El sistema vestibular está diseñado para obtener información sobre la postura y el movimiento, para lo cual es capaz de medir la aceleración lineal y angular de la cabeza a través de un dispositivo formado por cinco órganos sensoriales presentes en el oído interno (laberinto membranoso o vestibular).
Las funciones del SV se pueden resumir en tres:
- Proporcionar información a los órganos centrales del equilibrio del movimiento propio y del entorno para tener la sensación y conciencia de la orientación espacial del cuerpo en cada momento, lo que permite orientarse y desplazarse adecuadamente en el espacio.
- Desencadenar los oportunos reflejos osteomusculares necesario para el mantenimiento del equilibrio y la estabilidad corporal, tanto en reposo como en movimiento.
- Desencadenar movimiento cefálicos para la orientación de la cabeza en el espacio con el fin de estabilizar las imágenes retinianas mediante el reflejo vestibuloocular, lo que permite mantener la mirada fija en un punto del espacio cuando la cabeza se mueve.
Estas tres funciones del SV no son imprescindibles para el mantenimiento del equilibrio.
Para realizar estas funciones el sistema vestibular dispone de dos elementos básicos: los receptores sensoriales vestibulares con sus cc ciliadas y un arco neuronal de tres neuronas. Este arco neuronal está constituido por una neurona eferente bipolar, un procesador central interneuronal constituido por los núcleos vestibulares y los efectores que son las neuronas oculomotoras y los músculos.
Percepción de la sensación de orientación espacial.
Para ser consciente de la orientación espacial en un determinado momento es necesario percibir y recibir información de la orientación espacial adoptada por cada parte del cuerpo en cada momento y especialmente la de la cabeza.
La información de las aceleraciones que experimenta la cabeza son captadas por el SV y trasmitidas por el nervio vestibular principalmente hacia los núcleos vestibulares del bulbo raquídeo, desde donde se proyecta a los núcleos ventroposterior y ventrolateral del tálamo y de estos, a su vez, se proyectan en dos áreas corticales, la 2 y 3 de la corteza somatosensitiva primaria, en las que se genera una sensación subjetiva del propio movimiento del cuerpo y del entorno externo. Otra parte de la información vestibular se trasmite directa e indirectamente (por conexión con los núcleos vestibulares) al cerebelo, donde converge en la región conocida funcionalmente como vestibulocerebelo.
Esta información del movimiento y de la posición espacial de la cabeza está proporcionan por los receptores laberínticos. Estos son estimulados por los movimientos cefálicos y por la fuerza continua de la gravedad. Las aceleraciones lineales estimulan los otolitos maculares y las aceleraciones angulares y rotatorias los canales semicirculares. Los movimientos cefálicos y la fuerza de la gravedad son el estimulo captado por los receptores vestibulares y estos trasforman la fuerza mecánica del estimulo en impulsos nerviosos que son transmitidos, como hemos dicho, hasta los NV del tronco cerebral y de ellos, mediante conexiones nerviosas, al resto de los centros nerviosos relacionados con el equilibrio.
Los tres pares de canales, sistema canalicular, como se explicó en el tema 3.2ª, se disponen dentro del peñasco en tres planos del espacio, siendo aproximadamente perpendiculares entre sí, correspondiéndose cada par con un plano; con esta disposición al ser estimulados pueden captar cualquier movimiento cefálico, siendo el resultado de su estimulación la percepción de los movimientos cefálicos en las tres direcciones del espacio.
La combinación de los dos sistemas, canalicular y otolitico, asegura la captación de todos los movimientos cefálicos, señales que son trasmitidas al SNC para desarrollar una sensación subjetiva de conocimiento y consciencia de la posición de la cabeza en el espacio en cada momento (orientación espacial). Ahora bien, esta detección de los movimientos cefálicos tiene ciertas limitaciones, como más adelante se explica al exponer el funcionamiento de las corrientes endolinfáticas. Estas limitaciones hacen que ciertos desplazamientos pueden no ser bien detectados por el SV. Estos son;
- incapacidad de detectar movimientos lentos de la cabeza a velocidades constantes en el plano horizontal ya que sólo son estimulados por aceleraciones;
- incapacidad de distinguir una inclinación de la cabeza de una aceleración lineal de la misma.
Queremos hacer aquí una consideración sobre la importancia que tiene para el funcionamiento vestibular el equilibrio o balance entre el SV de ambos lados. Ambos laberintos se activan o se inhiben simultáneamente en condiciones fisiológicas, si un lado se activa el otro se inhibe: hay un balance correcto entre derecha e izquierda para conseguir una elaboración perfecta de datos que permita tener una orientación espacial correcta. Para realizar esta función de interbalance, el órgano más importante es el núcleo vestibular lateral pues es el origen del circuito de autorregulación vestibular homolateral. Ambos núcleos laterales y la formación reticular son los que intervienen en la simetría entre derecha e izquierda ayudados por otros circuitos de acoplamiento retrógrado que se superpone al anterior y que se localizan tanto en los propios NV, como en el cerebelo (ver en capítulo 02.3ª.05, vías eferentes vestibulares).
Esta función de percepción de la sensación de orientación espacial proporcionada por el SV no ha sido todavía lo suficientemente estudiada y raramente se evalúa en los protocolos clínicos.
Mantenimiento del equilibrio corporal en reposo y movimiento.
El SV contribuye al mantenimiento del equilibrio desencadenando dos tipos de reacciones musculoesqueléticas: reflejas y voluntarias
- Reflejas. Son reacciones musculoesqueléticas reflejas, que en coordinación con otros reflejos equilibratorios, ayudan a ajustar la postura y el equilibrio en respuesta a cambios en la posición y el movimiento de la cabeza y que son fundamentales para el mantenimiento del equilibrio.
El mantenimiento reflejo de la postura fundamental estática del cuerpo mediante una acción tonígena muscular permanente se realiza mediante la información aportada por el SV; esta sirve para que a través de mecanismos reflejos el cuerpo se mantenga permanente en equilibrio estático (tono muscular) y alineado con respecto al entorno: estabilidad postural en el espacio. Esta actividad tónica espontánea hace que medio cuerpo empuje al otro medio, consiguiendo así que esté en equilibrio. La estimulación tonígena, refleja y permanente, es ejercida por el SV sobre los músculos con acción antigravitatoria (extensores): reflejos vestíbulo-espinales.
Esta función se pone de manifiesto experimentalmente al hacer una laberintectomía en un animal, comprobando como se produce una hipotonía de ese lado, con lo cual el animal es incapaz de mantener el equilibrio, cayendo hacia el lado del laberinto destruido. Lo mismo ocurre en la especie humana, cuando se lesiona sólo el SV de un lado hay una disminución del tono muscular en el hemicuerpo del lado lesionado y el cuerpo cae hacia el lado hipotónico. Esta función es realizada, además de por el SV, por el reflejo miotático cuyo estímulo es el estiramiento muscular (circuitos propioceptivos intramedulares).
- Voluntarias. Las acciones musculoesqueléticas voluntarias son esenciales para el mantenimiento del equilibrio en situaciones donde se requiere un mayor control y adaptabilidad. Hay un uso consciente del sistema vestibular: al hacerse conscientes las señales del sistema vestibular a nivel cerebral las personas voluntariamente pueden ajustar su posición y movimiento para mantener el equilibrio. Algunas formas de utilizar conscientemente el sistema vestibular para mantener el equilibrio incluyen:
Fijar la mirada: el mantener la vista en un punto fijo ayuda a proporcionar referencias visuales estables que pueden ayudar al sistema vestibular a mantener la orientación y el equilibrio.
Movimientos coordinados: al coordinar conscientemente los movimientos de la cabeza y el cuerpo, se puede facilitar la interpretación de las señales vestibulares y ayudar en la adaptación a cambios en la posición.
Práctica de ejercicios de equilibrio: realizar ejercicios específicos que desafíen el equilibrio puede ayudar a mejorar la coordinación entre el sistema vestibular y las respuestas musculares.
Entrenamiento sensorial: La exposición gradual a situaciones que desencadenan respuestas vestibulares, como movimientos rotatorios o cambios en la posición, puede ayudar a mejorar la capacidad de respuesta y adaptación del sistema vestibular.
Orientación de la cabeza y de las imágenes retinianas.
La orientación de la cabeza y la estabilización de la mirada son dos funciones complementarias y necesarias para que la información captada por el SV sea correcta y para que la imagen a percibir por el ojo se centre en la retina al realizar cualquier movimiento cefálico.
- Orientación de la cabeza en el espacio. El SV desencadena reflejos para que la cabeza vaya orientándose durante la marcha o en cualquier otra situación de movimiento cefálico. Estos reflejos permiten que la cabeza esté estable y orientada espacialmente con relación al cuerpo en cada situación. La cabeza está siempre orientada de forma que el plano de la mácula utricular esté lo más próximo a la horizontal. Además es necesario que la orientación de la cabeza sea correcta en todo momento para que la captación de ciertas informaciones puedan ser bien percibidas e interpretadas, como son la altitud y la velocidad lineal.
- Estabilización de la mirada: el estímulo vestibular producido tras un movimiento cefálico, informa al cerebro de la amplitud y dirección que ha de tener el correspondiente movimiento ocular corrector para mantener la imagen del campo visual estable en la retina. Esta función se realiza mediante el reflejo vestíbulo-ocular: RVO. En la estabilización de la mirada, aunque sea de forma secundaria, también colabora el cuello por medio del RCO.
Como hemos dicho, ambos SV funcionan equilibradamente de forma que cuando uno se estimula, el otro se inhibe. Si la cabeza se mueve hacia un lado, el laberinto homolateral es el que se estimula, y el contralateral, el que se inhibe; el lado homolateral al movimiento predomina sobre el contralateral.
Aunque parezca contradictorio, la participación del SV en el equilibrio no es tan importante como podría deducirse de lo expuesto. Así, por ejemplo, los sordomudos con ambos laberintos anulados mantienen un perfecto equilibrio; en la tabes dorsal en que está anulada la información propioceptiva, al cerrar los ojos el enfermo se derrumba a pesar de tener unos laberintos posteriores perfectamente conservados.
Ahora bien, aunque la participación del SV en el equilibrio sea poca, su responsabilidad en el desequilibrio puede ser mucha.
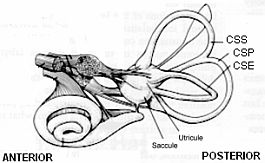
El aparato vestibular de cada lado posee cinco receptores vestibulares que se diferencian en dos grupos anatomofuncionales:
- Canales semicirculares: sus receptores son las crestas ampulares.
- Sistema otolítico o macular: máculas acústicas del utrículo y el sáculo.
Su función es ser estimulados por la acción de cualquier fuerza que actúe en un determinado momento sobre el organismo. Los diferentes tipos de fuerzas que ejercen su acción sobre el organismo lo hacen de diferente forma, según su naturaleza y dirección imprime un tipo diferente de movimiento. Para que una fuerza pueda ser estimulante no puede estar contrarrestada por otra similar de dirección contraria.
Cada receptor vestibular tiene un diseño especializado para la captación de un tipo de fuerza o movimiento con unas determinadas características (naturaleza y dirección de la misma), y según dicha fuerza estimule un determinado receptor el resultado sensorial del estímulo será también diferente.
La gravedad.
El cuerpo está sometido a la fuerza permanente de la gravedad terrestre. Para contrarrestar esta fuerza y no caer, el organismo tiene un sistema antigravitacional que funcionalmente corresponde al SV. Este dispone de gravirreceptores que son estimulados por la gravedad y un conjunto de palancas osteomusculares a las que envía los oportunos estímulos para darlos respuesta contrarrestando la fuerza gravitacional, permitiendo que el cuerpo venza la gravedad y no caiga.
Los gravirreceptores del SV son las máculas del sistema otolítico cuya composición es de un elevado peso específico lo que las hace ser susceptibles a la acción permanente de la gravedad a la que se halla sometido el cuerpo. Este estímulo es constante, salvo en circunstancias especiales que impulsan el cuerpo en dirección contraria al centro gravitacional terrestre igualando o anulando la fuerza de la gravedad y en los estados de ingravidez.
La detección precisa de la gravedad por parte del SV permite a nuestro cerebro ajustar constantemente las respuestas musculares y mantener el equilibrio, incluso en diferentes posiciones y movimientos. Ahora bien, en el sistema antigravitacional o de equilibramiento intervienen no sólo el SV, sino los tres elementos sensoriales de la triada de oritación témporo-espacial coordinadamente. Estas órdenes se plasman en una serie de reacciones que se diferencian en varios tipos: de enderezamiento, de equilibración y estáticas. Cuando las palancas osteomusculares fallan sobreviene la caída vertical por impulso gravitacional, es decir dirigida al centro terrestre.
El movimiento.
Otra de las fuerzas que estimula el SV es el movimiento, pero para comprender como actúa el movimiento para estimular al SV hay que diferenciar el movimiento cuando es uniforme y el movimiento cuando no es uniforme, pudiendo ser este último de aceleración o desaceleración. Esto es importante, ya que no es el movimiento en sí el que estimula a los receptores vestibulares, sino el movimiento no es uniforme, es decir, las aceleraciones ya sean en sentido positivo o negativo. El movimiento uniforme no estimula el laberinto.
- Aceleración lineal.
¿Las fuerzas de dirección lineal cómo estimulan al SV? Al iniciarse un movimiento lineal o rectilíneo, se produce una aceleración positiva hasta que la velocidad se hace constante o uniforme y en ese momento deja de ser estimulante. Los movimientos cefálicos lineales, ya sean verticales u horizontales, estimularán al sistema otolítico.
- Aceleración centrípeta.
Es la aceleración que experimenta un cuerpo cuando realiza o es sometido a un movimiento circular uniforme. Esta se produce por fuerzas de dirección elíptica, parabólica o circular. En virtud del principio de inercia estas fuerzas originan fuerza centrífuga. La fuerza centrífuga se asemeja mucho en su mecanismo de acción a la fuerza gravitatoria.
- Aceleración angular.
Es la que experimenta un cuerpo que se desplaza con un movimiento acelerado describiendo una trayectoria circula, no lineal. Esta viene expresada en grados. Esta aceleración ocupa un lugar preferente en los mecanismos de estimulación del SV, estimulando preferentemente a los receptores de los CS.
- Aceleración Coriolis, efecto Coriolis o fuerza Coriolis.
Es una "fuerza" adicional perpendicular al movimiento que experimenta cualquier objeto o cuerpo en movimiento rotatorio desviándolo de su recorrido y que da lugar a una trayectoria curva.
Estimulantes inadecuados.
Además de los estímulos descritos a los que podemos denominar fisiológicos, ya que son necesarios para el mantenimiento del equilibrio corporal, existen otros tipos de estimulantes de los receptores vestibulares a los que se les denomina inadecuados, o no fisiológicos. Estos, si bien son capaces de estimular el epitelio sensorial del SV, no lo hacen de forma funcional para el mantenimiento del equilibrio corporal.
Estos estímulos son el térmico, el galvánico y el sonoro.
Existe una concepción clásica de que los estímulos adecuados para las máculas utrículo-saculares son las aceleraciones linéales y los cambios posicionales de la cabeza, mientras que las crestas ampulares de los CS son estimulados solamente por las aceleraciones angulares. Esto hoy en día se sigue aceptando pero con ciertas restricciones como iremos viendo más abajo.
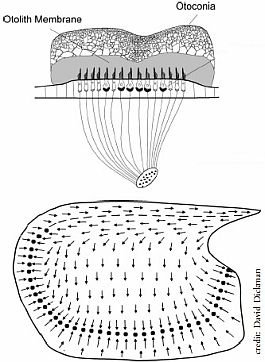
Las funciones del útrículo y del sáculo en el equilibrio corporal son todavía ampliamente discutidas, y sobre todo, la del sáculo no es conocida con exactitud. La fisiología funcional de cada máculas es diferente. Los estímulos más eficaces detectados por las máculas otolíticas son las aceleraciones o desaceleraciones lineales en los 3 planos del espacio.
Mecanismo de estimulación de las máculas.
La membrana otolítica con sus otolitos, junto con la membrana reticular con sus cc ciliadas, constituyen el sistema mecanoreceptor de las máculas.
Estos mecanorreceptores son estimulados por:
- La gravedad. Como ya dijimos los otolitos tienen mayor densidad que el líquido endolinfático que los rodea y al ser atraídos por la fuerza de la gravedad se desplazan dentro de la membrana otolítica.
- Las aceleraciones lineales desplazan a los otolitos por la misma razón que la gravedad, es decir, por la densidad relativa y la inercia de estos cristales de calcio en comparación con el líquido endolinfático circundante.
Como se dijo en la histología de los receptores, las cc ciliadas de cada mácula tiene sus cilios orientados en una dirección contrapuesta a cada lado de la estriola, esto hace que el desplazamiento de la membrana otolítica en una dirección tiene como consecuencia fisiológica una estimulación opuesta sobre el conjunto de las cc ciliadas a cada lado de la estriola. Al moverse la membrana otolítica en una dirección se estimulan la mitad de las cc cicliadas, aquellas en las que los esterocilios se inclinan hacia el kinocilio, no estimulándose las cc de la otra parte de la estriola. Los dos órganos sensoriales, utricular y sacular, están situados perpendicularmente entre ellos (ángulo recto de la mácula utricular respecto a la sacular). El estímulo más efectivo para estos receptores, aunque no el único, es la aceleración lineal realizada en un plano paralelo a cada mácula, comportándose como acelerómetros lineales e inclinómetros.
Teniendo en cuenta la orientación diferente de las máculas y la distribución celular de dirección opuesta, marcada por la estriola, las cc ciliadas están situadas en diferentes ángulos, lo que permite a las máculas tener una sensibilidad multidireccional para captar y responder a movimientos en los tres planos del espacio.
La membrana otolítica de las máculas tiene una densidad mayor que la endolinfa que las rodea, como dijimos al explicar la estimulación gravitatoria, esto permite que pueda desplazarse sobre el neuroepitelio sensorial. Por tanto el mecanismo excitatorio de estos receptores es el desplazamiento de la membrana otolítica sobre la capa celular que esta fija, adherida a la pared de utrículo y sáculo.
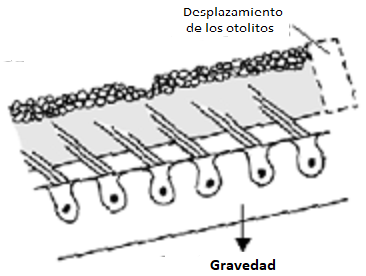 Como ya hemos dicho, la membrana otolítica es estimulada no sólo por las aceleraciones sino además por la fuerza de la gravedad especialmente al producirse cambios en la dirección de la gravedad de la tierra con respecto a la membrana. Así al producirse una inclinación de la cabeza el peso de la membrana otolítica hace que esta se deslice produciendo una fuerza sobre las cc ciliadas subyacentes. Al ser la gravedad un factor estimulador permanente hace que las máculas tengan una actividad en reposo constante.
Como ya hemos dicho, la membrana otolítica es estimulada no sólo por las aceleraciones sino además por la fuerza de la gravedad especialmente al producirse cambios en la dirección de la gravedad de la tierra con respecto a la membrana. Así al producirse una inclinación de la cabeza el peso de la membrana otolítica hace que esta se deslice produciendo una fuerza sobre las cc ciliadas subyacentes. Al ser la gravedad un factor estimulador permanente hace que las máculas tengan una actividad en reposo constante.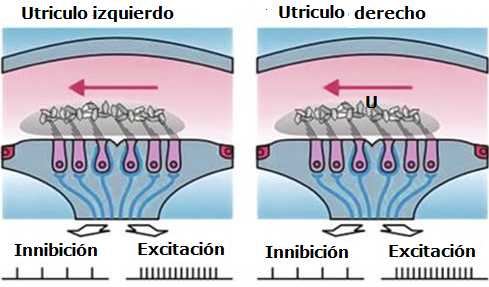 La mácula del utrículo, está colocada en el suelo, su orientación es horizontal en un plano más o menos frontal, su membrana otolítica captan sobre todo los desplazamientos lineales en el plano sagital (hacia delante y hacia atrás) y las inclinaciones de la cabeza tanto a los lados (lateralizaciones) como la flexo-extensión. La acción de ambas máculas utriculares es sinérgica para cualquier posición que adopte la cabeza girando sobre su eje bitemporal. Cuando la cabeza gira sobre su eje anteroposterior, la acción gravitatoria ejercida sobre las máculas utriculares no modifica la respuesta de éstas.
La mácula del utrículo, está colocada en el suelo, su orientación es horizontal en un plano más o menos frontal, su membrana otolítica captan sobre todo los desplazamientos lineales en el plano sagital (hacia delante y hacia atrás) y las inclinaciones de la cabeza tanto a los lados (lateralizaciones) como la flexo-extensión. La acción de ambas máculas utriculares es sinérgica para cualquier posición que adopte la cabeza girando sobre su eje bitemporal. Cuando la cabeza gira sobre su eje anteroposterior, la acción gravitatoria ejercida sobre las máculas utriculares no modifica la respuesta de éstas.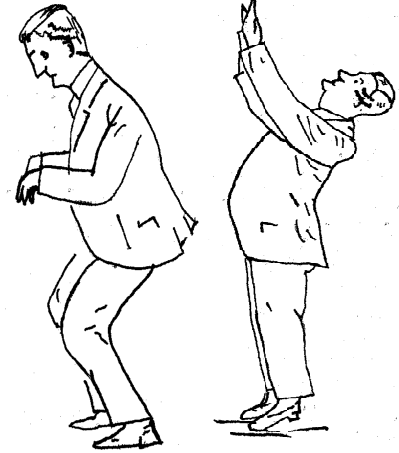 Cuando la cabeza está flexionada hacia adelante 30º, ambas máculas utriculares quedan en un plano totalmente horizontal ejerciendo una presión máxima sobre el órgano neurosensorial utricular siendo la posición en la que las máculas utriculares alcanza su máxima excitación. Cuando la cabeza se extiende 130º estas máculas toman una posición inversa a la anterior, que es la posición de mínima excitación para ellas. Esto se aprecia observando que cuando el sujeto flexiona la cabeza hacia adelante tiene una aptitud de flexión general, mientras que cuando la extiende adquiere una aptitud de extensión general.
Cuando la cabeza está flexionada hacia adelante 30º, ambas máculas utriculares quedan en un plano totalmente horizontal ejerciendo una presión máxima sobre el órgano neurosensorial utricular siendo la posición en la que las máculas utriculares alcanza su máxima excitación. Cuando la cabeza se extiende 130º estas máculas toman una posición inversa a la anterior, que es la posición de mínima excitación para ellas. Esto se aprecia observando que cuando el sujeto flexiona la cabeza hacia adelante tiene una aptitud de flexión general, mientras que cuando la extiende adquiere una aptitud de extensión general. Las máculas saculares, al estar situadas en un plano vertical (fronto-sagital), captan las aceleraciones de los movimientos cefálicos de ascenso y descenso (aceleraciones verticales) y por supuesto también las fuerzas gravitatorias. No son estimuladas por movimientos de aceleración lineal, sino por movimientos vibratorios de baja frecuencia. Al contrario de la utriculares que son sinérgicas sino que funcionan de forma antagónica entre ellas. cuando una está en hiperexcitación la otra está en hipoexcitación y viceversa.
Ambas máculas, aunque en menor medida, pueden ser estimuladas también por movimeintos de aceleraciones de traslación, centrífugos y centrípetos.
Funcionamiento del conjunto macular.
La acción de las máculas es sinérgica en cualquier posición que la cabeza adopte por giro sobre el eje bitemporal. Ambas máculas son agonistas al variar la posición alrededor del eje fronto-occipital. El estímulo gravitatorio por variaciones de posición sobre el plano horizontal no varia.
Los input-autput de los receptores maculares de cada lado, funcionan igual que en los CS, con la cabeza en reposo emiten una respuesta nerviosa igual de cada lado. El mecanismo fisiológico de la generación del impulso nervioso informador es el movimiento cefálico: al desplazarse la cabeza, las cc sensoriales se mueven también, pero la membrana otoconial al tener los cristales de carbonato cálcico es más densa que la endolinfa que la rodea, quedando retrasada a causa de la inercia. Este retraso es transmitido a través de la membrana gelatinosa produciendo la deflexión de los cilios que se encuentran inmersos en ella. El movimiento ciliar despolariza la cc desencadenando la estimulación celular, que aumenta o disminuye, según la dirección de la deflexión del cilio que depende de la dirección de la aceleración. Cuando se alcanza una velocidad constante, la membrana gelatinosa y la masa otoconial vuelven a su posición de reposo y no hay estimulación. Cuando la cabeza se retrasa, las cc sensoriales lo hacen igualmente al mismo tiempo, pero la otoconia tiene impulso y continúa moviéndose, quedando ligeramente adelantada. Este cambio relativo de posición se transmite de nuevo a los esterocilios por la membrana gelatinosa, pero la deflexión es ahora en sentido opuesto a la ocurrida durante la aceleración, por lo que la estimulación neuronal también se invierte.
Como hemos dicho, la gravedad es un estímulo constante para las membranas otoconiales lo que conlleva una actividad espontánea en reposo constante por parte de las cc ciliadas, está cifrada en 80 descargas por seg.
En la ingravidez se produce una desaferenciación de los órganos otolíticos, dejando de emitir descargas, pero esto no ocurre en los canales.
Esta actividad espontánea permanente del sistema macular cuando está en reposo, es decir, en ausencia de estímulos, se relaciona fisiológicamente con estas funciones:
▪ Contribuir al mantenimiento activo del tono muscular espontáneo, fundamental en reposo para el mantenimiento del equilibrio corporal. Diríamos que medio cuerpo empuja al otro medio consiguiendo el equilibrio, y si falla el laberinto de un lado, disminuye el tono muscular en el lado afecto predominado la otra mitad apareciendo desequilibrio. Por tanto, esta función es la que mantiene la bilateralidad y simetría que permite mantener el cuerpo en la línea media sin inclinarse hacia los lados.
▪ Permitir respuestas bidireccionales, o sea, que un estímulo es detectado no sólo en magnitud y duración, sino también en polaridad, como en el caso de aceleraciones positivas y negativas.
▪ Aumentar la sensibilidad del receptor, siendo su umbral de excitación más bajo.
Los neuroepitelios de los CS situados en las crestas ampulares son mecano-receptores preparados para ser estimulados por las aceleraciones angulares durante los desplazamientos de la cabeza; se comportan como acelerómetros angulares. Cada aceleración angular de la cabeza ejerce una acción específica sobre el sistema cúpulo-endolinfático del canal que se encuentre en el plano más próximo a la dirección de dicha aceleración. Los CS también son estimulados, aunque de forma muy secundaria, en menor intensidad, por las aceleraciones lineales, ahora bien, su estimulante especifico es la aceleración y deceleración angular.
La fisiología de la estimulación del neuroepitelio de los CS se explica por la teoría hidrodinámica: la fuerza estimuladora es el movimiento de la endolinfa. La corriente endolinfática produce una inclinación o un deslizamiento de la cúpula. Cuanto más enérgico es el movimiento mayor es la fuerza hidrodinámica del estímulo.
La cúpula al tener la misma densidad que la endolinfa, no se desplaza bajo el efecto de la fuerza de la gravedad, o por movimientos de la cabeza que simplemente modifican su línea de acción, como una inclinación de la cabeza, sino que se desplaza sólo por movimientos de aceleración angular que se producen con la rotación de la cabeza. Cada canal se estimula de forma optima durante el movimiento en torno a un eje que forme 90º con respecto al plano en el que está situado.
La fisiología y fisiopatología de los CS comenzó a conocerse con las investigaciones de Flourens (1830) y Carl Ewald (1889). Estos autores fueron los primeros en explicar la existencia de una relación clara entre los planos de los tres canales, la dirección del flujo endolinfático y la dirección de los movimientos de la cabeza y de los ojos. Sistematizaron cuales eran las respuestas musculares y nistágmicas a los estímulos ampulares mediante sus famosas leyes sobre la función canalicular. Estos conceptos hoy en día son contestados, ya que tienen poco de fisiológicos:
Ley de Jean Pierre Flourens: las respuestas musculares o nistágmicas se producen en el mismo plano del conducto estimulado. La destrucción de un conducto provoca movimientos, desviaciones corporales y oculares en el plano de ese conducto.
- Primera ley de Ewald: las respuestas musculares y nistágmicas (manifestaciones tónica) tienen la misma dirección que la corriente endolinfática que las provoca.
- Según la segunda ley de Ewald: en los canales horizontales las corrientes ampulípetas serían excitadoras y las ampulífugas inhibidoras. La excitación es más efectiva que la inhibición para cambiar la actividad laberíntica.
- Tercera ley de Ewald: en los CS verticales se invierte la segunda ley, es decir, es más activa la corriente ampulífuga que la ampulípeta.
De las leyes de Ewald se pueden deducir estos conceptos para comprender la fisiología del SV:
- Que existe una polarización de los CS.
- Que el factor de estimulación canalicular está constituido por las aceleraciones angulares.
- Que esta polarización de los CS se debe a como están organizados los cilios de las cc sensoriales, estas cc están polarizadas y el eje morfológico de polaridad, denominado vector de polarización, pasa por un manojo de esterocilios más cortos y por un kinocilio.
- Que la orientación del flujo endolinfático con relación a este eje de polarización determina el nivel de la actividad recogida a nivel de las aferencias vestibulares: las corrientes ampulípetas son excitadora y las ampulífigas son inhibidoras.
- Que la inclinación del esterocilio en dirección hacia el kinocilio (cinetocilio) se produce una activación de la cc, aumentando las descargas en las fibras eferentes, es decir una excitación; a la inversa, si la inclinación del esterocilio es contraria al kinocilio la cc se inhibe.
- Que los kinocilios están más próximos al utrículo en las crestas de los CSH y la inversa en los CSA y CSP, como demuestra la segunda ley de Edwald.
Si bien el movimiento, al producir corrientes endolinfáticas, es el estímulo fisiológico de estos receptores, también pueden ser estimulados al provocarse corrientes endolinfáticas mediante estímulos no fisiológicos (estímulos no fisiológicos) como el térmico, el galvánico, el sonoro (fenómeno de Tullio), los cambios de presión endolinfática (signo de la fístula de Lucae) y los cambios químicos en la endolinfa. Al estudiar las pruebas de exploración vestibular complementarías analizaremos estos fenómenos.
¿Cómo se estimulan los CS?
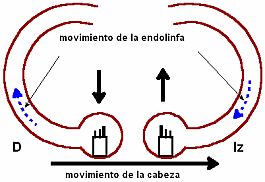 Cuando la cabeza realiza un movimiento, los canales se mueven al unísono con ella en la misma dirección. Al moverse el canal se desplazan igualmente las crestas ampulares, junto con sus cc ciliadas, como si estuviesen firmemente adheridas al hueso.
Cuando la cabeza realiza un movimiento, los canales se mueven al unísono con ella en la misma dirección. Al moverse el canal se desplazan igualmente las crestas ampulares, junto con sus cc ciliadas, como si estuviesen firmemente adheridas al hueso. Al realizar la cabeza un movimiento angular, el líquido endolinfático del canal no sigue la misma dirección de movimiento que el canal sino la opuesta. Esto es así porque al moverse el canal la columna de endolinfa dentro del mismo tiende a permanecer inmóvil por su propia inercia, es decir se opone a cambiar su estado de reposo ante la aceleración del movimiento de la cabeza. Al moverse el canal y no moverse el líquido, hay un desplazamiento del líquido a lo largo del canal semicircular en sentido-dirección opuesto al movimiento del giro cefálico: corriente endolinfática de inercia. Esta corriente endolinfática, que se origina retrasada al movimiento del canal, da como resultado una diferencia de presión a ambos lados de la cúpula, lo que hace que ésta se desplace.
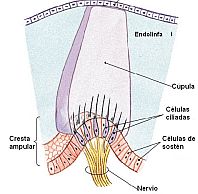 Como se dijo en la anatomía del laberinto membranoso, la cresta, cc sensoriales y la cúpula forman un diafragma que cierra herméticamente todo el perímetro del conducto a la altura de la ampolla. Este diafragma es elástico, siendo más móvil en su parte central que en sus extremos. En el interior de este diafragma la cúpula actúa a modo de válvula pudiendo someterse a movimientos de flexión sobre el eje de la cresta o bien deslizarse sobre ella dentro de la ampolla en ambas direcciones sin fricciones manifiestas. Una corriente endolinfática que se genere en su interior producirá una inclinación o desplazamiento de la cúpula. En ese momento la cúpula es desplazada, arrastrada, o presionada ligeramente por la corriente endolinfática y en la misma dirección de la corriente, pues sigue fielmente los movimientos de ésta, ya que tiene la misma densidad que la endolinfa. Al desplazarse la cúpula moviliza a los cilios de las cc neurosensoriales siendo estimulados.
Como se dijo en la anatomía del laberinto membranoso, la cresta, cc sensoriales y la cúpula forman un diafragma que cierra herméticamente todo el perímetro del conducto a la altura de la ampolla. Este diafragma es elástico, siendo más móvil en su parte central que en sus extremos. En el interior de este diafragma la cúpula actúa a modo de válvula pudiendo someterse a movimientos de flexión sobre el eje de la cresta o bien deslizarse sobre ella dentro de la ampolla en ambas direcciones sin fricciones manifiestas. Una corriente endolinfática que se genere en su interior producirá una inclinación o desplazamiento de la cúpula. En ese momento la cúpula es desplazada, arrastrada, o presionada ligeramente por la corriente endolinfática y en la misma dirección de la corriente, pues sigue fielmente los movimientos de ésta, ya que tiene la misma densidad que la endolinfa. Al desplazarse la cúpula moviliza a los cilios de las cc neurosensoriales siendo estimulados. La forma de desplazarse la cúpula es pivotando sobre la cresta ampular como un péndulo que posee una fuerte amortiguación, debido a que se encuentra inmerso en un medio líquido. Esta amortiguación pendular del sistema cúpulo-endolinfático impide la permanencia de un estímulo más tiempo del necesario para producir las correspondientes reacciones y además permite responder con gran sensibilidad a estímulos de corta duración. Los estímulos angulares tienen una magnitud mínima, tanto en cuanto al tiempo como a la intensidad y, en consecuencia la latencia de las respuestas a los estímulos es de muy corta duración.
Este mecanismo hidrodinámico que produce el desplazamiento de la cúpula estimulando los receptores ampulares está totalmente demostrado y admitido. Pero el mecanismo hidrodinámico no es el único mecanismo estimulatorio posible, ya que no es imprescindible la existencia de liquido en el interior del canal para que la cúpula se incline por el efecto de una aceleración angular, la propia inercia del sistema daría lugar, sin la necesidad de una fuerza hidrodinámica, a la inclinación de la cúpula sobre el neuroepitelio.
En los movimientos habituales de la cabeza la corriente endolinfática producida es muy pequeña, por lo que los movimientos de la cúpula son igualmente mínimos, así para un movimiento de cabeza de una velocidad angular de +/800º/seg la cúpula se desvía menos de un grado.
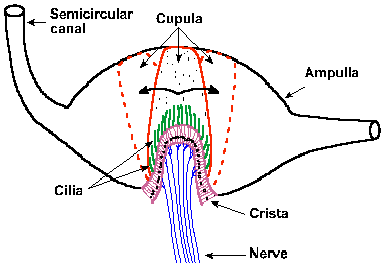 Al moverse el canal con las crestas ampulares, junto con sus cc ciliadas, los cilios, firmes en su base, son inclinados por el desplazamiento o torsión de la cúpula, curvándose. Recordemos que los cilios de las cc vestibulares son móviles, a diferencia de los cilios de las cc cocleares que son rígidos. La inclinación de los cilios movidos por la corriente endolinfática es el desencadenante de la estimulación de las cc sensoriales mediante su despolarización, modificando su actividad de reposo y formándose un potencial de acción. Estos fenómenos se producen en milisegundos siendo una característica de los receptores vestibulares que entre la detección del movimiento por el receptor y su percepción, es decir, el tiempo de latencia es muy corto, por esto los reflejos que desencadenan son muy rápidos.
Al moverse el canal con las crestas ampulares, junto con sus cc ciliadas, los cilios, firmes en su base, son inclinados por el desplazamiento o torsión de la cúpula, curvándose. Recordemos que los cilios de las cc vestibulares son móviles, a diferencia de los cilios de las cc cocleares que son rígidos. La inclinación de los cilios movidos por la corriente endolinfática es el desencadenante de la estimulación de las cc sensoriales mediante su despolarización, modificando su actividad de reposo y formándose un potencial de acción. Estos fenómenos se producen en milisegundos siendo una característica de los receptores vestibulares que entre la detección del movimiento por el receptor y su percepción, es decir, el tiempo de latencia es muy corto, por esto los reflejos que desencadenan son muy rápidos. Debido a la estrechez de los canales (0,4 mm), el fluido endolinfático sólo puede moverse longitudinalmente en una dirección a lo largo de su cavidad cilíndrica y la dirección del flujo será en una dirección en el canal de un lado y en la dirección opuesta en su homólogo del otro lado. Estos movimientos se denominan ampulípetos o utriculípetos y ampulífugos o utriculífugos según su dirección.
Con respecto al estímulo, cuando la aceleración va en aumento, la cúpula se desplaza cada vez más. Cuando la velocidad del movimiento cefálico es constante y uniforme, al no haber aceleraciones ni deceleraciones desaparece la inercia y el conjunto endolinfa-cúpula se mueve al unísono con el conducto, a la misma velocidad y en el mismo sentido, por lo que no hay corriente endolinfática y cesa la estimulación, ya que los cilios regresan a su posición vertical de reposo y no son estimulados. Cuando la aceleración cesa (deceleración) la cabeza y el conducto se detienen, pero la endolinfa continúa moviéndose durante un breve período de tiempo por la inercia, desplazando la cúpula en sentido contrario, originando la deflexión inversa de los cilios y en consecuencia una respuesta neural inversa.
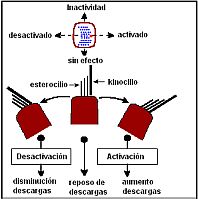 Estimulación de las células ciliadas.
Estimulación de las células ciliadas. La orientación del kinocilio en los receptores vestibulares es determinante para la fisiología vestibular. En los CS los kinocilios no están orientados en la misma dirección: en el CSH el kinocilio se sitúa en la cc mirando hacia el utrículo y en el CSP y CSA lo están en dirección opuesta. Por tanto, la estimulación resultante para cada CS será diferente según el sentido del giro.
En el CSH una corriente utriculípeta desplaza los esterocilios en dirección al kinocilio y provoca una aumento de información y cuando la corriente es utriculífuga produce el efecto contrario. Esto mismo se puede expresar así: una inclinación-deflexión de la cúpula hacia la ampolla produce estimulación de ese canal y una deflexión de la cúpula en dirección contraria a la ampolla hace disminuir los estímulos emitidos por el neuroepitelio de ese canal. El CSH y el utrículo orientan su acción en el plano horizontal, mientras que el CSP, el CSA y el sáculo lo hacen en un plano vertical. Es decir, que en el CSH la corriente ampulípeta es estimulante y en los verticales la corriente ampulífuga es la estimulante. En el CSP y CSA los desplazamientos ampulípetos de la endolinfa desplazan los cilios de las cc ciliadas en dirección opuesta la kinocilio generando estímulos inhibitorios, mientras que los desplazamientos ampulífugos generan estímulos excitatorios.
Funcionamiento del conjunto canalicular.
La velocidad de la corriente endolinfática es proporcional a la velocidad del movimiento cefálico, a más velocidad del movimiento más estímulo. Al girar la cabeza el laberinto de cada lado se mueve a una velocidad de aceleración distinta, el canal situado en la parte exterior del giro se desplaza a mayor velocidad que el situado interiormente, esto produce una corriente en los líquidos del laberinto situado en la parte externa del giro más intensa que en el situado internamente. Por tanto, se producirá un mayor número de descargas en el canal situado exteriormente que en el situado interiormente al giro cefálico.
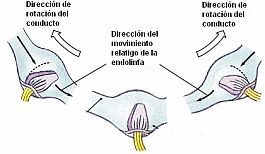 Este sistema se comporta fisiológicamente como una balanza, estando ambos lados organizados en un puch-pull o flip-flop, es decir, en inervación recíproca: así, un desplazamiento cupular utriculípeto en el CSH produce un aumento de la frecuencia de descargas de un lado y asocia un desplazamiento cupular utriculífugo en el CSH del otro lado con la consiguiente disminución de descargas. Cuando un lado es estimulado, el otro queda sometido a una inhibición en la producción de señales de tipo compensatorio. Cuando la cabeza está en reposo, los canales de cada lado que se hallan en el mismo plano, envían desde ambos vestíbulos descargas tónicas de la misma intensidad al tronco cerebral. Cuando la cabeza gira a una nueva posición según un determinado plano, la descarga tónica de uno de los canales en ese plano empezará a disminuir, a la vez que la del canal del lado opuesto empezará a aumentar. Cuando la cabeza rota en dirección opuesta ocurrirá lo contrario. Esta variación del "input" vestibular emitido para cada lado hacia el troncoencéfalo, al ser integrada por el SNC, se interpreta como giro.
Este sistema se comporta fisiológicamente como una balanza, estando ambos lados organizados en un puch-pull o flip-flop, es decir, en inervación recíproca: así, un desplazamiento cupular utriculípeto en el CSH produce un aumento de la frecuencia de descargas de un lado y asocia un desplazamiento cupular utriculífugo en el CSH del otro lado con la consiguiente disminución de descargas. Cuando un lado es estimulado, el otro queda sometido a una inhibición en la producción de señales de tipo compensatorio. Cuando la cabeza está en reposo, los canales de cada lado que se hallan en el mismo plano, envían desde ambos vestíbulos descargas tónicas de la misma intensidad al tronco cerebral. Cuando la cabeza gira a una nueva posición según un determinado plano, la descarga tónica de uno de los canales en ese plano empezará a disminuir, a la vez que la del canal del lado opuesto empezará a aumentar. Cuando la cabeza rota en dirección opuesta ocurrirá lo contrario. Esta variación del "input" vestibular emitido para cada lado hacia el troncoencéfalo, al ser integrada por el SNC, se interpreta como giro. Esta diferencia de actividad entre ambos laberintos es la señal que genera la respuesta vestibular. Con movimientos de la cabeza muy rápidos la actividad bioeléctrica espontánea en el CSH inhibido puede llegar a la desaparición, 0 impulsos nerviosos/seg., mientras en el CSH excitado puede aumentar en proporción a la intensidad del estímulo, pudiendo llegar a 400 impulsos/seg. Por tanto, el umbral para la percepción del movimiento por el aparato vestibular en el hombre es muy bajo, ya que somos capaces de percibir una aceleración de 0.1 grado/s2, esto es, el equivalente al giro de una silla que tarde unos 90 s en realizar una vuelta completa.
Los conducto CSH, al encontrase ambos en el mismo plano son sinérgicos. Respecto a los CSP y CSA se relacionan de la siguiente manera: EL CSP derecho se encuentra en el mimo plano que el CSA izquierdo y viceversa, el CSP izquierdo se encuentra en el plano del CSP derecho.
En términos generales se puede afirmar que el mecanismo mediante el cual un agente externo ejerce su acción sobre estos receptores es la deformación o inclinación de los cilios del neuroepitelio vestibular en diferentes direcciones.
Mecanismo de estimulación celular.
Las cc ciliadas de los neuroepitelios del SV son las encargadas de transformar la energía de los estímulos mecánicos en energía biológica, produciendo mensajes nerviosos.
Para que una fuerza sea eficaz como estimulo para las cc ciliadas tiene que actuar paralelamente a la superficie apical de la misma; una fuerza aplicada perpendicularmente a la superficie apical de la cc (fuerza de compresión), o en sentido lateral, no es eficaz como estímulo para estas cc; el estímulo es máximo cuando la fuerza se dirige a lo largo de un eje que divide el haz de esterocilios y pasa a través del kinocilio, es decir, longitudinalmente.
Polaridad de las células ciliadas.
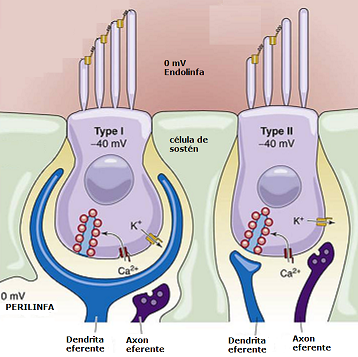
Existe una diferencia de voltaje entre el interior de la cc y el fluido que la rodea llamado potencial transmembranoso, que está originado por las diferentes concentraciones de iones a ambos lados de la membrana.
El potencial de reposo de las cc neurosenosriales es negativo (-80 mV) debido a que hay una mayor concentración de K y menor de Na y Cl en el espacio intracelular que en el extracelular.
Este potencial transmembranoso da lugar a un flujo continuo de corriente, debido sobre todo al K, desde el interior de la cc a través de sus paredes laterales hacia el espacio extracelular y desde allí hacia la membrana celular apical en contacto con la endolinfa.
Los ápices de las cc ciliadas están bañados por endolinfa. Las bases de estas cc están bañadas por la perilinfa que está fuera del laberinto membranoso. Por lo tanto, el ápice y la base de las cc ciliadas tienen ambientes eléctricos y químicos diferentes.
La endolinfa que rodea la cc ciliada es eléctricamente altamente positiva, la diferencia entre ambos potenciales intra y extracelular es de 135 mV. En este estado la cc de las máculas tienen una actividad bioeléctrica espontánea que, como hemos dicho, es de 80 descargas/seg. La estimulación de las cc ciliadas por inflexión de los cilios aumenta o disminuye su actividad, produciendo una respuesta excitatoria o inhibitoria, según el sentido o dirección del desplazamiento de los cilios.
Cambios en la polaridad celular.
 La membrana de las cc ciliadas es selectivamente permeable a la entrada y salida de determinados iones. Esta permeabilidad selectiva se realiza mediante la apertura y cierre de canales exclusivos para determinados iones, principalmente Na+, K+ y Cl++.
La membrana de las cc ciliadas es selectivamente permeable a la entrada y salida de determinados iones. Esta permeabilidad selectiva se realiza mediante la apertura y cierre de canales exclusivos para determinados iones, principalmente Na+, K+ y Cl++. La actividad de las cc ciliadas en uno u otro sentido viene determinada por su polarización morfofuncional, estando ésta relacionada con su organización ciliar, más concretamente con la orientación de los esterocilios con relación al kinocilio (o cinetocilio). La polaridad de la cc en cada momento viene determinada por la situación del quinocilio. La energía mecánica del flujo endolinfático que produce el desplazamiento o inclinación de los esterocilios en dirección hacia el quinocilio, produce un cambio en la resistencia de la membrana celular que da lugar a una apertura de los canales de K y como consecuencia una despolarización de la membrana celular y una reacción en cadena, disminuye la resistencia eléctrica entrando iones de Ca++ y salida de K++ en la cc, esta nueva concentración de iones a ambos lados de la membrana celular produce un cambio en la polaridad celular y su potencial se hace relativamente positivo y como consecuencia se produce una excitación celular positiva, que producirá la liberación de las aminas neurotransmisoras y la consecuente descarga de las fibras eferentes en la base de la cc. El quinocilio no es el responsable directo de la mecanotraducción, por lo que su manipulación no modifica el potencial de membrana celular. El quinocilio lo que hace es provocar el movimiento de los esterocilios. Es el desplazamiento de los esterocilios lo que produce la apertura de los canales iónicos de K. Para que estos canales se abran es necesaria la presencia de Ca.
La variación de voltaje producida en la vecindad de las cc por esta corriente de flujo se conoce como potencial microfónico de los órganos receptores. En contraste con los potenciales de acción de los nervios, los microfónicos de las cc ciliadas de los receptores no tienen periodo refractario y son muy resistentes a la anoxia.
Cuanto más grandes sean las desviaciones de los cilios hacia el kinocilio más se abrirán los canales de K y mayor será la despolarización, habrá mayor entrada de Ca a través de los canales de lo que dependerá el cambio de voltaje, mayor liberación de neurotransmisores y una mayor frecuencia de potenciales de acción en la fibra del nervio vestibular conectada a esa cc ciliada.
La deflexión de los cilios en la dirección opuesta, cuando las inclinaciones de los esterocilios son en dirección opuesta al kinocilio, disminuye la tensión en los enlaces de la punta, cerrando los canales de K a los que están unidos, esto hiperpolariza la cc ciliada y disminuye tanto la entrada de Ca como la liberación de neurotransmisores con la consiguiente disminución de descargas nerviosas o inhibición de las mismas. .
En los movimientos ciliares se encuentran implicadas la actina, miosina y tropomiosina que se encuentran en la base de los cílios.
Biología de las células ciliadas.
La corriente eléctrica asociada a los potenciales microfónicos en las sinapsis de las cc ciliadas interactúa con los neurotransmisores.
La excitación electrobiológica de las cc sensoriales se inicia con la síntesis de GABA que se deposita en las vesiculas sinápticas para ser liberada a la hendidura sináptica. El GABA es captado por los receptores de la membrana postsinática. Este proceso está modulado por los neurotramisores glutamato y sustancia P, situados a nivel del ápex de los cálices nerviosos aferentes, y mediante la liberación de GABA y acetilcolina en las terminaciones nerviosas eferentes. Por tanto la acetilcolina actúa como neurotransmisor eferente. Los neurotransmisores generan el potencial de acción postsináptico a través de la neuronas primarias aferentes.
Los potenciales así generados con directamente proporcionales al grado de inflexión de los kinocilios. A mayor movimiento del quinocilio, mayor cambio en los fenómenos bioeléctricos.
La orientación de las cc ciliadas de cada una de las tres crestas no es igual, como se puede deducir de la segunda ley de Ewald. En los CSH los quinocilios están polarizadas mirando hacia el utrículo: polarización ampulípeta. En los conductos semicirculares verticales la polarización es inversa: polarización ampulífuga.
La polarización morfofuncional u organización ciliar es distinta en las maculas del utrículo y del sáculo. Mientras en el utrículo el quinocilio se encuentra orientado hacia la estriola, en el sáculo se encuentra en la porción mas alejada a la misma. Por tanto, la polarización de la mácula utricular y sacular son opuestas. De todas formas en las máculas la direccionalidad ciliar no está tan bien definida como en las crestas y su patrón de respuesta es complejo.
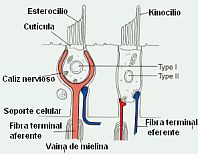
Las cc ciliadas en su polo basal sináptico entran en contacto con las fibras eferentes que transmiten la información hacia el nervio vestibular. Estas conexiones sinápticas son diferentes para los dos tipos de cc sensoriales de Wersall, cc tipo I y tipo II. Las cc tipo I hacen sinapsis solamente con una axón de grueso diámetro y que pertenece a una neurona dotada de actividad fásica; en las de tipo II la sinapsis se realiza con un axón que es de menor diámetro y que recoge la estimulacion procedente de la sinapsis con muchas cc y este axón corresponde a una neurona de actividad tónica. En este polo basal de las cc sensoriales nos encontramos también con fibras aferentes, llegadas de origen central, que tienen una función de control, modulando la sensibilidad dinámica de estas cc receptoras.
Las fibras nerviosas aferentes nacidas de los receptores están constituidas por los cilindroejes de neuronas vestibulares de primer orden, cuyo cuerpo celular se localiza en el ganglio de Scarpa. Estas neuronas están recibiendo información constantemente de las cc sensoriales, por lo que poseen una actividad denominada de reposo o actividad basal. Las neuronas del nervio vestibular mantienen un potencial de membrana típicamente en el rango de -70 a -80 milivoltios (mV). Cuando esta en actividad puede alcanzar una frecuencia de potenciales de acción de hasta aproximadamente 400 potenciales de acción por segundo.
En el utrículo y en el sáculo, cada cc sensorial se ve inhibida en un sentido y excitada en el otro. Hay un aumento de la frecuencia de los potenciales de acción en respuesta a una aceleración lineal en la dirección en la que las cc ciliadas están alineadas. Este aumento en la frecuencia de los potenciales de acción se interpreta en el cerebro como una indicación de movimiento o aceleración en esa dirección.
Una desaceleración o una aceleración en dirección opuesta a la orientación de las cc ciliadas produce una disminución en la frecuencia de los potenciales de acción.
Las neuronas del ganglio vestibular en reposos descargan espontáneamente en torno a los 100 potenciales de acción por segundo. Estas son aferentes que envían información sobre los movimiento cefálicos y eferentes que envían señales desde los órganos centrales del equilibrio al sistema vestibular para modular su sensibilidad.
El ganglio vestibular, no tiene diferente "tipos" de neuronas en sí mismo, sin embargo, las neuronas canaliculares y maculares se diferencian funcionalmente en varias categorías según se producen sus descargas en reposo y tras estimulación.
Las respuestas a las estimulaciones canaliculares diferencian fisiológicamente cuatro tipos de neuronas:
- Tipo I: son las más numerosas y entran en actividad al producirse aceleraciones angulares en dirección hacia su soma y su actividad es inhibida por la rotación opuesta. La actividad de descarga de estas neuronas vestibulares sigue un código bidireccional de las informaciones unidireccionales codificadas por los receptores laberínticos.
- Tipo II: estas neuronas tienen un patrón de respuesta inverso al de las neuronas de tipo I: son activadas por aceleraciones angulares de la cabeza en dirección opuesta a su soma e inhibidas por las rotaciones inversas. Estas neuronas forman además parte del sistema inhibidor intercomisural que comunica los NV de ambos lados.
- Tipo III y Tipo IV: son unas pocas neuronas que aumentan o disminuyen la frecuencia de sus descargas independientemente del sentido de la rotación de la cabeza.
Algunos autores hacen otra diferenciación funcional de las neuronas del ganglio vestibular en función del ritmo de sus descargas espontáneas en dos tipos:
- Neuronas regulares: son las que emiten descargas espontáneas de forma regular, sus fibras son de pequeño diámetro con una velocidad de conducción lenta; inervan una gran parte de la periferia del neuroepitelio sensorial de los receptores vestibulares. Otros autores las denominan tónicas.
- Neuronas irregulares son las que tienen sus axones de diámetro más grueso y su velocidad entro del neuroepitelio de los receptores vestibulares. Otros autores las denominan fásicas.de conducción es más elevada; son las principales neuronas aferentes; sus axones de estas neuronas están recubiertos con mielina, lo que aumenta la velocidad de conducción de los impulsos nerviosos; . inervan una zona más pequeña localizada en el c
Las respuestas a las estimulaciones maculares diferencian cuatro tipo de neuronas:
- Neuronas tipo alfa: suponen dos tercio de las neuronas del ganglio, por tanto constituyen las principales aferentes de las máculas. Tienen un gran cuerpo celular con un axón grueso y mielinizado. Son excitadas por la inclinación lateral ipsilateral de la cabeza y son inhibidas por la inclinación lateral contralateral. Funcionalmente en su gran mayoría su destino final es la región cervical.
- Neuronas de tipo beta: al igual que las anteriores tienen un cuerpo celular grande con un axón grueso y mielinizado. Su comportamiento fisiológico es el opuesto a las neuronas de tipo alfa.
- Neuronas de tipo gamma y delta: se activan e inhiben independientemente de la orientación que tenga la estimulación macular. Funcionalmente el destino final de las neuronas gamma es la región lumbar. Algunos autores no reconocen estos tipos de neuronas.
Las inclinaciones de la cabeza en el plano sagital siguen el mismo patrón que las del lateral.
Los NV integran las señales procedentes del nervio vestibular, la médula espinal, el cerebelo y el sistema visual, y se proyectan a diferentes centros: los núcleos oculomotores, los centros reticulares y medulares dedicados al movimiento esquelético, las regiones vestibulares del cerebelo (flóculo, nódulo, paraflóculo ventral y úvula) y el tálamo. Además cada núcleo vestibular se proyecta a otros núcleos vestibulares homolaterales y contralaterales.
Por tanto, el complejo de los NV además de constituir el primer centro de sinapsis de las vías vestibulares, no son un simple centro de conexión sino que constituyen un centro de integración multisensorial y motora, recibiendo aferencias espinales somato-sensoriales, visuales, auditivas, cerebelosas, corticales y vestibulares contralaterales. En ellos converge la información de la triada sensorial del equilibrio: vestibular, visual y propioceptiva. Integrando esta información elaboran una primera representación interna y tridimensional de los movimientos de la cabeza y el tronco en el espacio. Esta representación es fundamental para el mantenimiento de la estabilización de la mirada y la postura corporal y es el origen de la percepción del movimiento propio.
Los cuerpos neuronales de la segunda neurona que forman los NV tienen una actividad espontánea en reposo, que se modifica como respuesta a estímulos de aceleración angular cefálica.
Funcionalmente sus neuronas se diferencian en dos tipos:
- Tipo I: son las neuronas más numerosas, monosinápticas, su actividad aumenta con las rotaciones cefálicas ipsilaterales y disminuye por las rotaciones contralaterales. Funcionalmente pueden ser tónicas y cinéticas. Las tónicas mantienen una actividad espontánea que las hace estar descargando impulsos continuamente, y responden a estímulos por débiles que estos sean, ya que tienen un umbral muy alto. Las cinéticas (o fásicas) no tienen actividad espontánea, sólo se estimulan cuando llega un impulso directo desde el órgano vestibular periférico, siendo sensibles sólo a estímulos fuertes a los que responden inmediatamente, ya que tienen un umbral muy bajo.
- Tipo II: estas neuronas polisinápticas presentan un patrón fisiológico contrario a las de tipo I, disminuye su actividad espontánea ante estímulos ipsilaterales y aumenta ante estímulos contralaterales. Estas neuronas no son estimuladas directamente por las aferencias laberínticas primarias como las de tipo I, sino que lo son a través de las conexiones intervestibulares por las neuronas tipo I contralaterales. Las neuronas tipo II, tras ser estimuladas por las de tipo I contralaterales, tiene una función inhibitoria de las neuronas tipo I de su mismo lado. De esta forma los NV de un lado pueden controlar las rotaciones de la cabeza en ambas direcciones.
Ambos tipos neuronales tienen una capacidad de saturación cuando los estímulos son prolongados e intensos.
Los NV reciben información sobre la actividad nerviosa de ambos laberintos y efectúan un análisis comparativo de esta información bilateral, mediante el cual se evalúa el movimiento cefálico captado, según que la actividad aportada por cada lado sea igual o diferente.
Cuando los impulsos nerviosos que llegan a los núcleos de ambos lados son iguales, la actividad de los mismos es homogénea y entonces estos emiten impulsos nervioso homogéneos. Ahora bien, en cuanto se produce una diferencia de intensidad en los impulsos recibidos de cada lado, ésta se traduce, a su vez, en una diferente actividad en las cc nucleares de cada lado que comienzan a emitir estímulos por sus vías eferentes.
Es de interés resaltar la función del núcleo de Deiters. Este núcleo forma parte del sistema motor extrapiramidal y es un eslabón clave en los reflejos del equilibrio, ya que va a actuar a modo de compuerta, dejando pasar unos u otros impulsos reflejos, según las necesidades del momento.
Fisiología de cada núcleo vestibular.
Los NV se diferencian por su citoestructura en función de una segregación funcional, si bien en la clínica se consideran como un todo funcional.
Núcleo medial o de Schwalbe:
Sus neuronas son fundamentalmente excitadoras. Sus funciones fisiológicas son:
- Da origen al fascículo vestíbuloespinal medial ejerciendo un importante papel en los movimientos cervicales. Tiene un papel importante en la coordinación de los movimientos cervicales y oculares, envían fibras, a través del fascículo longitudinal medial, rostralmente hasta los centros oculomotores.
- Desempeña un papel crucial en el control de los movimientos oculares reflejos, especialmente aquellos relacionados con la fijación visual y la estabilización de la mirada durante los movimientos de la cabeza. Esto se logra mediante las fibras enviadas a través del fascículo longitudinal medial, rostralmente, hasta los centros oculomotores, actuando en coordinación con otras estructuras involucradas en el control del movimiento ocular, como es el núcleo superior que también está implicado en los reflejos que controlan la mirada.
- Tiene también conexiones cerebelosas que desempeñan un papel importante en la regulación y coordinación de los movimientos corporales, especialmente en relación con el equilibrio y la postura.
Núcleo lateral o de Deiters:
Recibe información de todos los receptores vestibulares.
Da origen a los axones que forman el fascículo vestibuloespinal lateral para las motoneuronas de la médula espinal. La función de este fascículo es favorecer el tono de la musculatura extensora tanto de forma directa como indirecta mediante las interneuronas: reflejos vestibuloespinales.. Es de interés resaltar que este núcleo forma parte del sistema motor extrapiramidal y es un eslabón clave en los reflejos del equilibrio, ya que va a actuar a modo de compuerta, dejando pasar unos u otros impulsos reflejos, según las necesidades del momento.
Núcleo superior o de Bechterew:
Sus neuronas se proyectan sobre los núcleos oculomotores a través del fascículo longitudinal ascendente y su principal función es regular el reflejo vestibuloocular. Sus neuronas son fundamentalmente inhibidoras.
Núcleo inferior o de Roller:
Se proyecta bilateralmente sobre el cerebelo vestibular y sobre el núcleo fastigial por el fascículo vestibuloespinal medial. Su función principal es la de coordinar con los núcleos contralaterales. La información que transportan el núcleo superior e inferior es diferente.
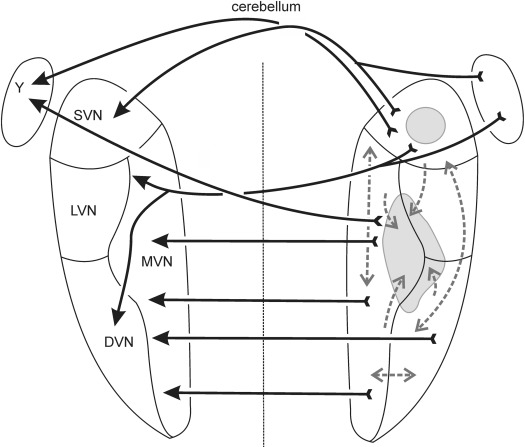
No existen conexiones aferentes vestibulares de la primera neurona que crucen la línea media; todas las conexiones entre los núcleos de un laberinto con los núcleos del lado opuesto se realiza a través de la segunda neurona o mediante conexiones más altas.
La funcionalidad de las conexiones entre los núcleos de cada lado es que cada neurona de un lado puede ejercer una influencia negativa depresora o excitadora sobre su neurona homóloga situada en los núcleos del otro lado, se trata fundamentalmente de impulsos inhibidores.
Estas conexiones permiten que en circunstancias normales, a pesar de que la actividad vestibular durante el movimiento cefálico sea asimétrica, no sea interpretada por el SNC como disbalance entre ambos lados, sino como movimiento cefálico bien de giro, o de lateralización, etc.. La actividad durante el movimiento está controlada por el lado excitado, que además de transmitir a las vías vestibulares la excitación recibida, mediante estas conexiones, coordinar la diferente actividad entre ambos lados. Así, un paciente con pérdida unilateral de la función vestibular carece de dicho mecanismo de interrelación entre ambos lados y los movimientos cefálicos muy rápidos hacia el lado de la lesión suelen producir inestabilidad corporal y ocular.
Estas conexiones desempeñan un papel clave en la integración de la información sensorial relacionada con el equilibrio y la orientación espacial.
Estas son las funciones más importantes asociadas con las conexiones intervestibulares:
- Integración de la información vestibular: permiten la integración de la información proveniente de los dos lados. Esto es esencial para obtener una imagen completa y precisa de la posición de la cabeza y el cuerpo en el espacio.
- Coordinación de la actividad bilateral importante para mantener el equilibrio y la postura de manera simétrica y para responder eficientemente a los cambios en la orientación y el movimiento. Gracias a su actividad reciproca realizada por estas conexiones contribuyen a la generación de respuestas compensatorias en todo el cuerpo que realizan ajustes corporales para contrarrestar los cambios en la posición y el movimiento de la cabeza.
- Estabilización de la mirada (fijación visual): permiten una coordinación eficiente de los movimientos oculares en ambos ojos. contribuyendo a la estabilización del campo visual durante movimientos de la cabeza. Esto es necesario para mantener la fijación visual durante los movimientos de la cabeza y para estabilizar la imagen visual durante la locomoción y otros movimientos corporales.
- Coordinación con otros sistemas sensoriales: la información vestibular se integra con la información visual y propioceptiva a través de las conexiones intervestibulares y otras vías neurales. Esta integración sensorial es fundamental para la percepción completa y precisa de la posición y el movimiento del cuerpo en el entorno.
- Respuestas de adaptación: desempeñan un papel en las respuestas de adaptación del sistema vestibular. Esto implica ajustes continuos y automáticos en la respuesta vestibular para adaptarse a cambios en el entorno o en las condiciones de movimiento.